Effect of Cooking and Reconstitution Methods on the Loss of Bioactive Compounds in Pigmented and Unpigmented Potatoes ()
1. Introduction
Population-based epidemiological studies have stressed the important role of diet and lifestyle in the emergence of many degenerative chronic diseases, such as cancers and cardiovascular diseases, in both developed and developing countries [1] . If a global health goal is to increase the amounts of vitamins, minerals or phytonutrients consumed in the diet, then a sensible approach is to further enhance the nutritional content of highly consumed crops. The nutritional value of the potato (Solanum tuberosum L.) is worth considering with regard to the high consumption of potatoes in the world and the body of evidence showing the relationship between diet and human diseases [1] [2] . Potato is currently the fourth most important crop worldwide after rice, wheat and maize with about 324 million tonnes production in 2010 [3] . The annual diet of an average global citizen in the first decade of the 21st century included about 33 kg of potatoes. In many developed countries, potato represents a secondary staple crop, with an average per capita consumption of around 75 kg∙year−1 in 2007 [4] . In developing countries, its consumption is around 20 kg∙year−1 per capita but rapid expansion over the past few decades has occurred in southern and eastern Asia [4] . In the Andes of South America, the potato consumption can reach 250 kg∙year−1 per capita. In these regions, potato constitutes the main staple crop, and most households cultivate 10-12 varieties in order to reduce their vulnerability to environmental conditions [5] .
Potato is an exceptionally high-yielding carbohydrate-rich crop and much of the world’s production is occupied by modern yellow-and white-fleshed potatoes. Other notable features are a high-quality protein, a significant level of vitamin C [6] [7] [8] that contributes substantially to the Recommended Daily Allowance [8] , also certain minerals such as potassium, phosphorus, and magnesium [5] . Beyond these basic nutrients, less well known are significant amounts of bioactive compounds (including phenolics and carotenoids) in potato which are potent antioxidants [9] [10] .
Promising routes for enhancing the quality of staple crops include biotechnologies and conventional plant breeding. Although some efforts to improve potato carotenoid [11] or phenolic contents [12] are currently made through transgenic approaches, natural variations in the potato germplasm may also offer nutritionally interesting potato genotypes. Of particular interest are the native Andean potato landraces (pigmented potatoes). Andean potato landraces are extremely diverse as compared to modern potato varieties, ranging from diploids to pentaploids; a wide variability in tuber shape, flesh and skin color, and flavour; in storage and cooking quality [10] [12] [13] and, so far, are largely unexplored from nutritional point of view.
One study has shown that antioxidant values (ORAC) for red-fleshed types reached as high as 300%, while for purple-fleshed types reached 250% of the white-fleshed genotypes [14] . Lechman et al. [7] found high variability in anthocyanin contents and antioxidant capacity (ABTS, FRAP and DPPH test) among 30 diverse coloured genotypes. The purple-fleshed clones were among the top. Lewis et al. [15] found that cultivated potato tuber peel contained 2000 - 5000 µg per g FW phenolic acids and 200 - 300 µg of flavonoids. Purple- and red-fleshed tubers contained three to four times the concentration of phenolic acids and had twice the flavonoid concentration of white-fleshed genotypes. Lewis et al. [15] also found much higher concentrations of anthocyanins in certain genotypes extending up to 368 mg per 100 g FW in the purple-fleshed and up to 22 mg per 100 g FW in red-fleshed types. Concentrations were considerably higher in peel, approaching 900 mg in purple-fleshed and 500 mg in red-fleshed types per 100 g FW (of the peel alone).
Given that potato is not consumed fresh by humans, it is fundamental to measure the bioactive compounds after cooing. However, other than domestic cooking; there is little published information on the effects of commercial cooking/processing on its constituents (except vitamin C) particularly in different colour-fleshed potatoes. Physical and/or chemical (degradation or losses of sensitive nutrients) alterations are possible depending on methods used to process potatoes [6] [16] [17] and obtaining processed products that keep more of their original characteristics are one of the most tangible challenges in potato processing industry, because of their content of antioxidant metabolites which are readily prone to thermal degradation. If a substantial amount of potato bioactive components survive cooking, then introducing new genotypes with even higher amounts would be a logical approach to boosting dietary intake.
There are contradictory reports on the effects of cooking on bioactive compounds of potato in the literature. A number of studies have charted the loss of vitamin C during diverse cooking and processing steps. In a review, Brown [18] reported that 20% to 45% of vitamin C is lost due to diffusion into the water bath during blanching (an important part of pre-cooked French fry production) and processing of potato into flakes almost totally eradicated vitamin C. Other studies have shown that different cooking treatments reduce various phenolic compounds in potatoes [6] [16] [19] . One study examining the effects of cooking procedures on mature potatoes showed that baking in an oven at 212˚C for 45 min, boiling in water for 30 min, or microwaving at 218˚C for 30 min, decreased chlorogenic acid to 0%, 35%, and 55% of the original amount, respectively [20] .
In contrast, scientifically optimized microwaving (2 min 30 s at high in an 1100 W microwave), baking (375˚C for 30 min), boiling (18 min), steaming (15 min) and stir- frying in cold-pressed canola oil (18 min) of developmentally young (small sized) potatoes of three genotypes resulted in an increase in recoverable amounts of the total phenolics, chlorogenic acids, rutin, kaempferol-rutinose, vitamin C and antioxidant activities [8] . Other studies showed an increase in most of the individual phenolic compounds and antioxidant activity cooked by baking, frying and microwaving when compared to uncooked tuber samples [7] [8] . However, published data on the extent of variation with regard to phytonutrient contents in commercially processed potato products especially within the colour-fleshed potatoes are scarce.
Aims of this study were to quantify the bioactive components (ascorbate, total antioxidants, total phenolics and anthocyanin) of seven potato genotypes and selections and evaluate how the commercial processing methods and subsequent freezing and final reconstituted cooking methods affect their bioactive components.
2. Materials & Methods
2.1. Potato Genotypes
Seven potato genotypes were used in this study. Name, flesh colour, average tuber size and dry matter content of the genotypes are presented in Table 1 and Figure 1. Among those, Adirondack Red, Adirondack Blue and AR2009-10 were Agriculture and Agri-
![]()
Figure 1. Skin and flesh colour of the potato genotypes. Potato genotypes: Clockwise from the left; AR2009-10 (recently registered as “AAC Blue Steele”), Adirondack Red, Adirondack Blue, Anuschka, Congo, POROIPG22-1, and Russet Burbank.
![]()
Table 1. Name, flesh colour, average tuber size and dry matter content of the genotypes.
*Average weight ± SE (n = 28). **Mean dry matter content ± SE (n = 4).
Food Canada (AAFC) selections; POROIPG and “Anuschka” were McCain Foods Ltd. Canada selections; Congo and “Russet Burbank” were used as control. AR2009-10 has now been registered as a variety “AAC Blue Steele” in Canada.
2.2. Field Trials
The tubers were grown on the research farm of McCain Foods Ltd., 200 Greenfield Road, Greenfield, New Brunswick, Canada (46˚27.58'N, 67˚38.06'W). The soil of the trial plot was predominantly Caribou gravelly silt loam series; well drained with less than 5% slope on most of the farm; high in P and K; medium in Ca2+ and Mg2+; 2.3% - 2.9% organic matter and 5.5 - 6.8 pH.
On 25 May 2012, plots were staked off in the field and furrows opened and fertilizer (16-9-30 3B at 200 kg∙ha−1) banded using a modified pea planter. Cut seeds (~60 g in weight) of the seven genotypes were hand dropped at 30 - 40 cm spacing in 9 m long rows for each genotype. Each plot (9.0 m × 1.82 m = 16.38 m2) consisted of two rows at 91 cm apart. Seeds were covered using disks mounted to a tractor. The plots were arranged in a randomized complete block design with four replications. Hilling was completed once 20 days after planting using a single pass row forming hiller. Standard pest protection measures were taken with routinely sprayed fungicides (mainly to control late blight) and insecticides (mainly to control Colorado potato beetle and flea beetles). The plants received a total of 366 mm of precipitation during the 120-day growth period (since 25 May to 25 September 2012). Maximum and minimum daily temperatures during the growing period ranged between 10.6˚C to 31.3˚C and 0.8˚C to 19.8˚C respectively.
On 25 September 2012, plots were harvested with a two-row potato digger and picked up by hand. After grading, the tubers were stored at the McCain Food Research Farm storage at 13˚C and 90% RH. A portion of the representative tuber samples (around 25 tubers for each genotype and replicates) were shipped to AAFC, Kentville, Nova Scotia (NS) laboratory and portions of the rest tubers were processed at McCain’s commercial potato processing facility [on 20 Nov. 2012 (rep 1), 22 Nov. 2012 (rep 2), 4 Jan. 2013 (rep 3) and 16 Jan. 2013 (rep 4)]. Based on types of industrial products, defined methods for processing, cooking and reconstitution were selected. Processing methods were blanching and freezing; cooking methods were microwave and frying; reconstitution methods were oven baking and frying. Combinations of these methods were made, generating five types of samples: Blanching-freezing (BF), Blanching- freezing microwave (BFM), Blanching-frying-freezing (BFF), Blanching-frying-freezing oven (BFFO), and Blanching-frying-freezing fryer (BFFF) samples. Step by step potato processing at McCain Foods Ltd. has been presented in Figure 2. The processed samples were shipped (in dry ice) to AAFC, Kentville, NS. The samples were subsequently freeze-dried, ground and stored at −80˚C until further analysis.
2.3. Tuber Storage and Tissue Sampling
The refrigerated shipment from McCain Foods Ltd., NB, Canada of around 25 whole tubers for each genotype and replication were received at AAFC, Kentville, NS on 11 October 2012. Tubers were stored immediately at 13˚C and at 90% RH. After four weeks, the storage room temperature was reduced to 12˚C. Thereafter, the temperature was reduced by 1˚C every two weeks till it reached to 9˚C and this temperature was maintained till the end of the study. The humidity remained always the same (90% RH) until the end of the study.
2.3.1. Initial Fresh (IF) Samples
Initial fresh samples were processed during ascorbate analysis on 22, 23, 24 and 25th of October 2012. For ascorbate analysis, seven whole tubers were randomly selected from each genotype and replicate. The tubers were washed under tap water, dried with Kim wipes, and weighed in a tarred bowl collectively on a weighing balance and tuber mass was recorded. Then two 1 cm thick coins were cut from the central portion from each
![]()
Figure 2. Step by step potato processing (at MaCain Foods Ltd.) followed for this study. Note: Samples and steps involved; BF = 1 - 9 & 15; BFM = 1 - 9 & 13 - 15; BFF = 1 - 13; BFFO and BFFM = 1 - 15.
of the seven tubers. The remaining portions of the tubers were put aside. The coins were cut again at the center portion to take two ~1 cm2 bars. The skin from both ends of each bar was removed. The 14 bars (from seven tubers) were chopped together into small pieces and only 5 g of these fresh samples were used for ascorbate analysis. Formoisture and dry matter content determination, ~20 g of the chopped samples were placed in an oven set to 85˚C for 72 hin pre-weighed closed paper bags. After cooling, the tissue weights were recorded. The percentage of moisture contents and dry matter were calculated as:

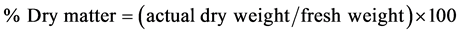
The remaining portions of the seven tubers were longitudinally cut into two halves. One half was discarded and the other halves from each portion were chopped together into small pieces as ascorbate samples but including the skin. These with skin samples and the left over (without skin) samples from sacorbate analysis were thoroughly mixed together, frozen in liquid nitrogen, freeze-dried and preserved at −80˚C which comprised initial fresh (IF) samples. Therefore, IF samples were actually a combination of with skin (~60%) and without skin (~40%) samples.
2.3.2. Fresh “with Skin” (FWS) and Fresh “No Skin” (FNS) Samples
These samples were processed on 9th, 10th, 11th and 22nd of January 2013, respectively, for Rep 1, 2, 3 and 4 (about 9 - 10 weeks after processing the initial fresh samples). For FWS and FNS samples, eight whole tubers from each genotype and rep were randomly chosen from the storage, washed and dried as described above. Each tuber was longitudinally cut into halves. Then one half was peeled using a standard kitchen peeler (at 1 - 2 mm depth) and chopped together. The other halves were chopped together with skin. The samples were separately frozen in liquid nitrogen, freeze-dried and preserved at −80˚C similarly as initial fresh samples.
The above sampling produced three groups of fresh samples for each genotype. After the industrial processed samples arrived, they were freeze-dried and stored at −80˚C until further analysis. Together, final samples for each genotype/selection were: I) Initial fresh (IF); II) Fresh with skin (FWS); III) Fresh no skin (FNS); IV) Blanching- freezing (BF); V) Blanching-freezing microwave (BFM);VI) Blanching-frying-freezing (BFF); VII) Blanching-frying-freezing oven (BFFO); and VIII) Blanching-frying-freez- ing fryer (BFFF).
2.4. Determination of Total and Reduced Ascorbate Contents
Ascorbate (as ascorbic acid, AsA) was extracted and analyzed using fresh potato tubers based on method developed by Bartoli et al. [21] with minor modifications. Approximately 5 g tissue from the finely chopped and mixed samples (mentioned above) was ground in 5 mL of 5% metaphosphoric acid using a pre-chilled ball mill (Teflon bowl with two zirconium balls) at 25 frequencies for 2 min.The homogenates were decanted into a 50-mL plastic centrifuge tube and another 10 mL of 5% metaphosphoric acid was used to wash the Teflon bowl and zirconium balls and decanted into the same 50 mL plastic centrifuge tube. The homogenates were centrifuged at 10,000 ×g for 15 min at 4˚C. The supernatant was removed to a cooled clean 50 mL plastic centrifuge tube and placed on ice. For reduced ascorbate determination, a 200 µL supernatant aliquot was combined with 1000 µL of 150 mM phosphate buffer (pH 7.4; 5 mM EDTA) and 200 µL of Milli-Q water (Millipore, Bedford, MA). For total ascorbate, the same procedure was followed except that 200 µL of 5 mM dithiothreitol was substituted for 200 µL of Milli- Qwater. The standards were prepared as per the reduced ascorbate determination, except that 200 µL of each standard was substituted for the sample aliquot. Samples and standards were incubated at room temperature for 15 min in the dark. O-phosphoric acid (100 µL) was added to each sample and standard to neutralize dithiothreitol and to acidify the solution for high performance liquid chromatography (HPLC) analysis. Samples were then filtered through 0.2 µm polyvinylidene fluoride filters (Chromatographic Specialties Inc., Canada) fitted to a glass syringe that was rinsed three times with methanol between samples. Reduced and total ascorbates were quantified with an isocratic HPLC (Waters Corp., Milford, MA) with a C18 guard and analytical column (Luna 5 µL 150 × 4.6 mm i.d., Phenomenex, Torrance, CA). Recoveries for ascorbate spiked samples were >95%.
2.5. Determination of Total Phenolics
Total phenolics were extracted and analyzed based on the method developed by Singleton and Rossi [22] and Folin and Ciocalteu [23] with some modifications. Powdered freeze-dried samples for each genotype and replicate were taken from −80˚C and 0.250 g of white/yellow or 0.125 g of pink/purple/blue potato tissue samples were weight out in 50 mL centrifuge tubes. A 10 mL of extraction solution (40% acetone, 40% methanol, 20% Milli-Q water and 0.1% formic acid) was added, vortexed and sonicatedat#15 for 30 sec and left under dim light for 30 min. The sample was then centrifuged at 10,000 × g for 15 minand the supernatant was transferred to clean 50 mL centrifuge tubes and kept in fridge until use. A second extraction was done similarly and the supernatant was added together to have a 20 mL volume. A 25 µL of standard or sample extract into 4 wells each of a 96-well microplate was used for activity assay at 750 nm on a Multiskan Spectrum microplate reader (Thermo Fisher Scientific, Ventaa, Finland) using 250 µL Milli-Q water, 50 µL Folin-Ciocalteu reagent and 12.5 µL saturated Na2CO3. Regression line was generated from the standard absorbance and used to quantitate the samples absorbance to obtain “mg∙gE/g DW” (mg∙gallic Acid Equivalent/g Dry Weight) values using the following formula:
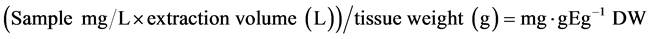
2.6. Determination of Total Antioxidant Capacity by ORAC
Total extraction (without separation into lipo/hydro fractions) was carried out under dim light. About 50 mg fine ground lyophilized potato tissue was weighed into 50 mL centrifuge tubes and placed on ice. Then 10 mL 70% acetone, 29.5% Milli-Q water and 0.5% acetic acid (AWA) was added to each sample and sonicated at #15 for 1 min with tube on ice. It was then centrifuged at 10,000 × g for 15 min and the supernatant was transferred to 25 mL volumetric. The extraction was repeated 2 times, each time the supernatant was transferred to the same volumetric. After the extraction, volume was brought to 25 mL in volumetric using AWA and kept on ice. For white and yellow tissues (for both fresh and processed samples), 300 µL supernatant was removed in an eppendorf tube and 600 µL AWA (room temp) was added (3 × dilution) and vortexed. For fresh pink/purple/blue tissues, 200 µL was removed in eppendorf tubes and 1000 µL of AWA (room temp) was added to tube (5 × dilution) and vortexed. For processed pink/purple/blue tissues, 300 µL was removed and 600 µL AWA (room temp) was added (3 × dilution) and vortexed. The samples were analyzed on a Fluoroskan Ascent FL 96-well microplate reader (Thermo Electron Corp., Vantaa, Finland) using 2,2’- azobis (2-amidinopropane) dihydrochloride (AAPH) as a peroxyl generator and 6- hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (Trolox) as a standard according to the method developed by Prior et al. [24] . The antioxidant capacity was expressed as TE∙g−1 DW.
2.7. Determination of Anthocyanin
Anthocyanins were determined based on the methods developed by Lee et al. [25] with some modifications. About 50 mg finely ground lyophilized potato tissue was weighed into 50 mL centrifuge tubes. A 10mL of extraction solution (40% acetone, 40% methanol, 20% Milli-Q water and 0.1% formic acid) was added, vortexed and sonicated at #15 for 30 sec and left under dim light for 1 h and 30 min but vortexed every 10 - 15 min. The sample was then centrifuged at 15,000 × g for 15 min then 8 mL of the supernatant was transferred into clean glass tubes and placed under flow of nitrogen. After drying, 2 mL of Milli-Q water was added, shaken well and transferred intopre-weighed small glass vials and placed for freeze-drying. After freeze-drying, weight of the vials were recorded again and 1 mL of Milli-Q water was added to each vial. Two 96-well microplates were used for each set of samples (sample + pH 1.0 buffer; and sample + pH 4.5 buffer) for assay at 700 nm and 520 nm on a “Multiskan Go” microplatereader (Thermo Fisher Scientific, Ventaa, Finland). Final volume in wells was 300 µL. The results were expressed as mg C3GE−100g FW (Cyanidin-3-glucoside equivalents).
2.8. Statistical Analysis
The treatments were arranged as a two-factor (potato genotypes and cooking/recanta- tion methods) factorial randomized complete block design with four replications. Data were subjected to analysis of variance using SAS (SAS Institute, Inc., Cary, NC). When F was significant at the P < 0.05 level, treatment means were compared and separated using the least significant difference test (LSD). Regression analyses were performed to investigate the relationships betweenphenolics vs antioxidant capacity; anothocyanins vs total antioxidant capacity; anthocyanins vs total phenolics; and total ascorbate vs total antioxidant capacity.
3. Results
3.1. Total, Reduced and Oxidized Ascorbates
Total ascorbate (AsA) contents significantly varied (P < 0.0001) between genotypes and ranged between 152 - 358 µg∙g−1 FW with the highest content in “Anuschka”, followed by Adirondack Blue (315 µg∙g−1 FW; Figure 3(a)). The lowest amount was determined in POROIPG22-1 (152 µg∙g−1 FW) followed by Adirondack Red (195 µg∙g−1 FW). Russet Burbank, Congo and AR2009-10 had similar and intermediate amounts of total ascorbates (~250 µg∙g−1 FW). Reduced ascorbate contents followed similar trends among the genotypes and on average of all genotypes was about 76% of the total ascorbate (i.e. ~24% oxidized ascorbate, Figure 3(a)). Regarding oxidized ascorbate, AR2009-10 had the highest amount (93 µg∙g−1 FW) followed by Congo (74 µg∙g−1 FW). Adirondack Red, POROIPG22-1, Anuschaka and Russet Burbank had the lowest amounts of oxidized ascorbates. Oxidized ascorbate content of Adirondack Blue was statistically similar with
![]() (a)
(a)![]() (b)
(b)
Figure 3. Total, reduced and oxidized ascorbate contents (a) and oxidized/reduced ascorbate ratio (b) of the seven potato genotypes (uncooked). The bars represent mean values of four replicates (±SE). Means with different letters (within a group) are significantly different at P < 0.05. The boxed value indicates percent of total ascorbate content.
Congo, Anuschka and Russet Burbank (Figure 3(a)). Regarding the ratio of oxidized/ reduced ascorbate, AR2009-10 had the highest oxidized/reduced ascorbate ratio followed by Congo. Anuschka had the lowest oxidized/reduced ascorbate ratio and other genotypes were statistically similar with either Anuschka or Congo (Figure 3(b)).
3.2. Total Phenolic Contents in Fresh Potatoes
Genotypic differences in total phenolic contents, effects of processing/reconstitution and interaction between genotype × processing were highly significant (P < 0.0001). Total phenolics contents in initial fresh (IF, 60% no skin, 40% with skin) samples ranged between 1.88 to 3.96 mg∙g−1 DW. Adirondack Blue, AR2009-10 and Adirondack Red had similar and significantly (P < 0.05) greater amounts compared to POROIPG22-1 and Congo. Anuschka and Russet Burbank had similar and the least mounts of total phenolics (Figure 4(a)).
![]() (a)
(a)![]() (b)
(b)
Figure 4. Total phenolics contents of the seven uncooked potato genotypes (a) and effect of cooking (average over all genotypes) on total phenolics (b). The bars represent mean values of four replicates (±SE). Bars with different letters within a group are significantly different at P < 0.05. The boxed values indicate average over all genotypes. IF = Initial fresh; FWS = Fresh, with skin; FNS = Fresh, no skin; BF = Blanching-freezing; BFM = Blanching-freezing microwave; BFF = Blanching-frying-freezing; BFFO = Blanching-frying-freezing oven; BFFF = Blanching-frying- freezing fryer.
Total phenolic contents in fresh-with skin (FWS) samples ranged between 1.67 to 4.52 mg∙g−1 DW (Figure 4(a)). In general, coloured potato genotypes (AR2009-10, Adirondack Blue, Adirondack Red, Congo and POROIPG22-1) had 1.7- to 2.5-fold greater phenolics than the yellowish/white genotypes (Anuschka and Russet Burbank). Adirondack Blue had the highest amount of phenolics (4.52 mg∙g−1 DW) and the lowest was in Russet Burbank (1.67 mg∙g−1 DW). However, the content in AR2009-10 was statistically similar with Adirondack Blue and Adirondack Red. POROIPG22-1, Congo, Anuschka and Russet Burbank respectively, in this order, had significantly (P < 0.05) lower amounts of total pehnolics compared to Adirondack Blue, AR2009-10 and Adirondack Red.
Peeling in general caused significant reduction in phenolic contents. Total phenolic contents in fresh-no skin (FNS) samples ranged between 1.34 to 3.87 mg∙g−1 DW (11% to 28% reductions compared to FWS samples) with the highest reduction in POROIPG22-1 (28%) followed by AR2009-10, Anuschka and Russet Burbank (~20% reduction). Reduction in other genotypes ranged between 11-14% (Figure 4(a)). On average over all genotypes, FWS samples had significantly higher amounts of total pehnolics (3.27 mg∙g−1 DW) compared to IF (3.05 mg∙g−1 DW) and IF samples had significantly greater total phenolics compared to FNS (2.71 mg∙g−1 DW) samples.
3.3. Effect of Processing/Cooking/Reconstitution on Total Phenolics
In this study, we aimed at investigating the effects of processing/cooking/reconstitu- tion, therefore, the data were averaged over all genotypes. It was found that fresh with skin (FWS) samples had the highest value at 3.27 mg∙g−1 DW total phonolics (Figure 4(b)). Initial fresh (IF) and fresh no skin (FNS) samples had, respectively, 7 and 18% lesser phenolics compared to FWS samples which were statistically significant (P < 0.05; Figure 4(b)). Cooking/reconstitution of the potatoes, blanching-freezing (BF) in particular, eliminated about 65% phenoics compared to FWS (and 58% compared to FNS). Further cooking/reconstitution processes (microwave, frying, oven and fryer) did not cause additional reductions in phenolic contents (Figure 4(b)).
3.4. Effect of Processing/Cooking/Reconstitution on Phenolics of Individual Genotype
Regarding individual genotypes, phenolic contents in IF and FWS samples did not differ significantly (P > 0.05) in most genotypes except for Adirondack Blue and POROIPG22-1. However, peeling (FNS samples) caused significant (P < 0.05) amount of phenolic losses (compared to FWS) in most genotypes except for Congo and Russet Burbank (Table 2(a)).
Blanching removed most of the pehnolics in all genotypes with little or no further reductions with further processing/reconstitution, except for Congo. Congo had kept significantly (P < 0.05) greater (~7%) amounts of phenolics in BF samples compared to other cooking/reconstitution methods (Table 2(a)). In general, genotypes with greater initial levels of phenolics (coloured genotypes) lost more (~65%) and genotypes those had less phenolics to start with (Anuschka and Russet Burbank), lost less (~50%) to cooking/reconstitution (Table 2(a)).
3.5. Anthocyanin Contents in Fresh Potatoes
All genotypes were analysed for anthocyanin contents, however, no anthocyanins were detected in Anuschka (yellowish flesh) and Russet Burbank (creamy white flesh). Therefore, anthocyanin data have been presented excluding Anuschka and Russet Burbank (Table 2(a), Figure 5). Genotypic differences in anthocyanin contents, effects of processing/reconstitution and interaction between genotype ×processing were highly significant (P < 0.0001). In initial fresh (IF) samples, AR2009-10 had the highest amount of anthocyanins followed by Adirondack Blue and Adirondack Red andPOROIGP22-1 had significantly lower anthocyanins compared to Adirondack Red, how-
(a) ![]() (b)
(b) ![]()
Table 2. (a) Effect of potato processing/cooking on total phenolics and anthocyanins of individual potato genotypes; (b) Effect of potato processing/cooking on total antioxidant capacity of individual potato genotypes.
Note: Means with different lower case letters in a column are significantly different at P < 0.05. IF = Initial fresh; FWS = Fresh, with skin; FNS = Fresh, no skin; BF = Blanching-freezing; BFM = Blanching-freezing microwave; BFF = Blanching-frying-freezing; BFFO = Blanching-frying-freezing oven; BFFF = Blanching-frying-freezing fryer.
ever, contents in Congo were the lowest (Figure 5(a)). Anthocyanin contents in fresh- with skin (FWS) samples ranged between 13.98 to 38.57 mg C3GE−100g FW (Figure 5(a)) with the highest amount in AR2009-10 (38.57 mg C3GE−100g FW) followed by Adirondack Blue (23.80 mg C3GE−100g FW). AdirondackRed, POROIGP22-1 and Congo had similar and the lowest amounts of anthocyanins (~14.5 mg C3GE−100g FW). Trends in fresh-no skin (FNS) samples were similar to that of FWS samples (Figure 5(a)). On average over all genotypes, FWS samples had the highest (21.24 mg
![]() (a)
(a)![]() (b)
(b)
Figure 5. Anthocyanin contents in five uncooked coloured-flesh potato genotypes (a) and effect of cooking (b) on anthocyanins (average over all genotypes). The bars represent mean values of four replicates (±SE). Bars with different letters within a group are significantly different at P < 0.05. The boxed values indicate average over all genotypes. IF = Initial fresh; FWS = Fresh, with skin; FNS = Fresh, no skin; BF = Blanching-freezing; BFM = Blanching-freezing microwave; BFF = Blanching-frying-freezing; BFFO = Blanching-frying-freezing oven; BFFF = Blanching-frying- freezing fryer.
C3GE−100g FW) anthocyanins. Initial fresh (IF) and FNS samples had, respectively, 10% and 19% lesser anthocyanins compared to FWS samples which were statistically significant (P < 0.05; Figure 5(b)).
3.6. Effect of Processing/Cooking/Reconstitution on Anthocyanin Contents
On average over all genotypes, cooking/reconstitution of the potatoes, blanching in particular (BF), eliminated about 81% the anthocyanins compared to FWS samples (and 77% compared to FNS). Further cooking/reconstitution processes (BFF, BFFO and BFFF), similarly and significantly (P < 0.05) promoted further reductions (~10% more) compared to BF samples (Figure 5(b)). Anthocyanins in BFM samples were statistically similar with the BF samples and also were not significantly different from the BFF, BFFO and BFFF samples.
3.7. Effect of Processing/Cooking/Reconstitution on Anthocyanins of Individual Genotype
Regarding individual genotypes, peeling (FNS) did not significantly (P > 0.05) reduce anthocyanins in other genotypes except for AR2009-10 and POROIPG22-1 (Table 2(a)). Regarding initial fresh (IF) samples, only AR2009-10 had significantly (P < 0.05) lesser amount of anthocyanins compared to FWS samples (Table 2(a)). Nevertheless, it possessed the highest amounts of anthocyanins in fresh samples compared to all the other four genotypes. Cooking/reconstitution processes, blanching in particular, greatly eliminated anthocyanins in all genotypes with no further reductions with further processing/reconstitution except for Adirondack Blue. Adirondack Bluehad significantly (P < 0.05) greater (~7%) amounts of anthocyanins in BF samples compared to other cooking/reconstitution methods (Table 2(a)).
3.8. Total Antioxidant Capacity of Fresh Potatoes
Genotypic differences in total antioxidant capacity, effects of processing/cooking/re- constitution and interaction between genotypes × processing were highly significant (P < 0.0001). In general, pigmented genotypes (AR2009-10, Adirondack Blue, Adirondack Red, Congo and POROIPG22-1) had 2- to 3-fold greater antioxidant capacities than the unpigmented genotypes (Anuschka and Russet Burbank; Figure 6(a)). Total antioxidant capacity in initial fresh (IF) samples ranged between 31 to 101 TE g−1 DW. Adirondack Red had greater antioxidant capacity compared to AR2009-10. However, antioxidant capacity of Adirondack Blue was not significantly (P > 0.05) different from Adirondack Red and AR2009-10. Congo and POROIPG22-1, in this order respectively, had significantly (P < 0.005) lower antioxidant capacity compared to Adirondack Red, AR2009-10 and Adirondack Blue. Russet Burbank and Anuschka had similar and significantly (P < 0.005) lower antioxidant capacity compared to all five other genotypes (Figure 6(a)).
Total antioxidant capacity in fresh-with skin (FWS) samples ranged between 35 to 110 TE∙g−1 DW. Adirondack Red and Adirondack Blue had similar and the highest antioxidant capacity followed by POROIPG22-1 and AR2009-10 (~95 TE∙g−1 DW).Congo, Russet Burbank and Anuschka, in this order, respectively had significantly (P < 0.05) lower antioxidant capacity compared to other genotypes (Figure 6(a)). Fresh-no skin (FNS) potatoes had significantly (P < 0.05) lesser antioxidant capacity compared to fresh-with skin (FWS) potatoes and ranged between 27 to 92 TE∙g−1 DW (Figure 6(a)). Maximum loses were recorded in POROIPG22-1 and Russet Burbank (~35%) and the least reductions were recorded in AR2009-10, Adirondack Blue and Adirondack Red (~16%). On average over all genotypes, total antioxidant capacity of FWS samples was 80 TE∙g−1 DW. About 21% antioxidant capacity was lost due to peeling. Initial fresh (IF) samples had about 13% lesser antioxidant capacity compared to FWS samples which were statistically significant (P < 0.05; Figure 6(a)).
![]() (a)
(a)![]() (b)
(b)
Figure 6. Total antioxidant capacity of seven uncooked potato genotypes (a) and effect of cooking (b) on total antioxidant capacity (average over all genotypes). The bars represent mean values of four replicates (±SE). Bars with different letters within a group are significantly different at P < 0.05. The boxed values indicate average over all genotypes. IF = Initial fresh; FWS = Fresh, with skin; FNS = Fresh, no skin; BF = Blanching-freezing; BFM = Blanching-freezing microwave; BFF = Blanching-frying-freezing; BFFO = Blanching-frying-freezing oven; BFFF = Blanching- frying-freezing fryer.
3.9. Effect of Processing/Cooking/Reconstitution on Total Antioxidant Capacity
On average over all genotypes, blanching alone (BF samples) reduced 58% antioxidant capacity compared to FWS samples (45% compared to FNS samples).Further cooking/ reconstitutions (BFF, BFFO and BFFF), also similarly and significantly (P < 0.05) reduced total antioxidant capacity (~12%) except for BFM processing (Figure 6(b)).
3.10. Effect of Processing/Cooking/Reconstitution on Antioxidant Capacity of Individual Genotype
Regarding individual genotypes, peeling (FNS) significantly (P < 0.05) reduced total antioxidant capacity in all genotypes except Anuschka. Total antioxidant capacity of IF samples were significantly (P < 0.05) lesser in most genotypes compared to FWS samples except AR2009-10, Congo and Anuschka (Table 2(b)).
Blanching greatly reduced total antioxidant capacity in all genotypes with some differences among them. Further cooking/reconstitution did not cause further reductions in antioxidant capacity except for Adirondack Red, Adirondack Blue and AR2009-10 (Table 2(b)). BF samples of these three genotypes had about 13% more antioxidant capacity compared to BFF, BFFO and BFFF samples. However, as contrary to the total phenolics, genotypes with greater initial levels(pigmented genotypes) and genotypes with lesser initial levels (Anuschka and Russet Burbank); similarly lost about 65% of the total antioxidant capacity to blanching and further cooking/reconstitution (Table 2(b)).
3.11. Bioactive Compounds Loses Due to Processing and Relationships between Total Phenolics, Anthocyanins, Total Antioxidant Capacity and Total Ascorbates
On average over all genotypes, ~65% of the total phenolics were lost during potato processing. Of which, ~18% was lost to peeling, 40% to blanching and ~7% was lost during further cooking/reconstitution (BFM, BFF, BFFW, BFFF). About 90% of the anthocyanins were lost during potato processing. Of which about 20% was lost to peeling, ~60% to blanching and ~10% was lost during further cooking/reconstitution. While about 70% of the total antioxidant capacity were lost during potato processing. Of which, about ~23% was lost to peeling, ~35% to blanching and ~12% was lost during further cooking/reconstitution (Figure 7(a)).
The results showed that there were highly significant (P<0.001) positive correlations between total phenolics and total antioxidant capacity (Figure 7(b)); anothocyanins and total antioxidant capacity (Figure 7(c)); and anthocyanins and total phenolics (Figure 7(d)). However, there were significant (P < 0.05) negative correlation between total ascorbate and total antioxidant capacity (Figure 7(e)).
4. Discussion
Total phenolics, anthocyanins and antioxidant capacity in fresh samples of five colour/pigmented and two unpigmented potato genotypes/selections were quantified and how commercial processing/reconstitution processes affect them were investigated in this study. Ascorbate profiles of the seven genotypes were also investigated using fresh tuber/tissue only (as available literatures show that there are little or no ascorbates in commercially processed potatoes). The results showed that the ascorbate profiles of the genotypes were not associated with a particular flesh colour/pigment. However, pigmented potatoes in general (with significant variations between them) contained higher levels of total pehnolics and anthocyanins compared to unpigmented genotypes. The pigmented potatoes also had higher antioxidant capacity in scavenging ORAC (oxygen radical performance capacity) radicals. The commercial processing/reconstitution processes, peeling and blanching in particular, appeared to have significant effects on phenolics, anthocyanins and antioxidant capacity as their levels were largely reduced in commercially processed/reconstituted products in all genotypes.
4.1. Potato Genotype Differences in Bioactive Compounds
The seven genotypes greatly varied in their contents of bioactive compounds. These
![]() (a)
(a)![]() (b) (c)
(b) (c)![]() (d) (e)
(d) (e)
Figure 7. Losses of bioactive compounds from potatoes during possessing/cooking (a); and relationships between total phenolic contents and total antioxidant capacity (b); anthocyanin contents and total antioxidant capacity (c); total anthocyanins and phenolic contents (d); total ascorbate content and total antioxidant capacity (e) of the seven potato genotypes. TA capacity = Total antioxidant capacity.
findings are explained by the genetic variations in these genotypes. Genotype differences in bioactive components are common in vegetables and fruits as has been seen in this study. The coloured potatoes had 1.7 to 2.5 times more the phenolics presumably due to the high anthocyanins in them and about 2 to 3 times more antioxidant capacity compared to unpigmented genotypes. These findings are in consistence with [5] [17] [26] and [27] who also reported greater amounts of bioactive compounds in coloured potatoes. No anthocyanins were detected in the unpigmented potatoes however; coloured genotypes had higher levels of anthocyanins. Brown [18] and Lewis et al. [15] also reported that outside of the center of origin of cultivated potato in the Andes of South America, it is rare to find varieties with anthocyanin pigments.
High antioxidant capacity in pigmented potatoes in our study can be explained by the presence of higher amounts of total phenolics and anothocyanins in them compared to unpigmented potato genotypes. Figures 6(b)-(d) also justify these even substantial altercations in their contents after cooking. Faller and Fialho [28] also reported positive correlations between polyphenols and antioxidant capacity in raw and cooked potatoes, carrots, onions, broccoli, and white cabbage. Lachman et al. [7] showed that total anthocyanins and individual anthocyanid corresponded to the antioxidant activity of various red and purple fleshed potatoes. Brown et al. [26] reported 330% greater antioxidant activity in a red fleshed breeding line of potatoes than the white fleshed potato breeding lines and varieties, as has been seen in this study. Brown et al. [26] also reported that vitamin C may also account for up to 13% of the total antioxidant capacity of potato. Mohdaly et al. [29] also reported high antioxidant capacity as strongly related to the presence of numerous phenolic and flavonoid compounds in various vegetables and fruits. Although we did not analyze carotenoids, coloured potatoes are rich in carotenoids and are also correlated with antioxidant activity in potato tubers [26] .
Under storage conditions, bioactive components of vegetables and fruits generally reduce over time. However, during the 9 - 10 weeks in storage (at 13˚C - 9˚C and 90% RH), potato tubers seemed did not lose phenolics or antioxidant capacity significantly. The lesser amounts of phenolics in initial fresh (IF) samples (compared to FWS) were due to the combination of “with” and “without skin” samples in IF samples explained in the methodology section.
4.2. Bioactive Compounds in Coloured Potatoes vs Berries
Nevertheless, weighing against other researches those used similar quantification methods as performed in the present study for different kind of berries [30] [31] , it was found that coloured potatoes had levels of phenoliocs, anthocyanins and antioxidant capacity that reached levels similar to those observed in strawberries and raspberries; and reached about 25% to those observed in blueberries. Ascorbate levels in coloured potatoes were similar to unpigmented genotypes, also similar to those observed in strawberries and raspberries; and about three times more than blueberries. Unlike berries, potatoes are served as part of a main course meal and, therefore, incorporation of pigmented flesh potatoes in meal can provide additional natural sources of potent antioxidants in the diet. Moreover, potatoes cost less to produce and result in higher yields that can also be economical source of raw material for extraction of anthocyanins. Along with carotenoids, anthocyanins are hydrophilic pigments responsible for the red, purple, and blue hues in most flowers, grasses, fruits, vegetables, and grains [5] . The association of coloring properties and potential health benefits in anthocyanins [32] makes them very attractive molecules for the food industry. Taken together, our results provide further evidence for the enhanced health beneficial components in pigmented potatoes. Mulinacci et al. [17] suggested that a portion (about 200 g raw) of these coulourful potato genotypes could consistently contribute to the daily intake of these functional compounds.
4.3. Ascorbate Contents in Coloured Potatoes
Ascorbate contents were not found to be associated with the tuber colour as Anuschka had the highest amount of ascorbate, while POROIPG22-1 had the lowest and “Russet Burbank” were somewhere in the middle with other coloured potatoes. However, AR2009-10 was recorded with the highest oxidized ascorbate contents and highest oxidized/reduced ascorbate ratio, followed by Congo (Figure 1(a) and Figure 1(b)) indicating less monodehy droascorbate reductase (MDHAR) and dehydroascorbate reductase (DHAR) activities in these genotypes, because these two enzymes are responsible for reducing the oxidated ascorbate to the reduced stage in order for its antioxidant functions..
Since total ascorbate is the sum of reduced ascorbate and oxidized ascorbate, the ascorbate pool in plants is maintained about 90% in reduced form with the remaining 10% present in the oxidized form [33] . The key aspects governing the ascorbate pool in plants are its biosynthesis, redox turnover, and degradation rates. The turnover occurs through a regenerative process. Reduced ascorbate is oxidized (by ascorbate oxidase, ascorbate peroxidase, etc. enzymes) to monodehydroascorbic acid (MDHA). MDHA is converted to reduced ascorbate through the activities of MDHAR ordispropotionates non-enzymatically to oxidized ascorbate. Oxidized ascorbate can be re-reduced back to reduced ascorbate directly through DHAR or undergoes irreversible hydrolysis to 2,3- diketogulonic acid which are toxic secondary metabolites such as oxalate and tartarate [34] [35] . Therefore, MDHAR and DHAR are important enzymes contributing maintaining an appropriate level of reduced ascorbate concentration in plant cells which seemed low in genotypes AR2009-10 and Congo compared to other genotypes.
However, the overall ascorbate pool in potato tubers in our study was found as 76% in reduced from and 24% in oxidized form (instead of 90:10). This could be due to the reduced MDHAR and DHAR activity in tuber tissues (sink) as compared to the photosynthetically active shoot/leaf (source) tissues. MDHAR and DHAR enzyme activity has been reported to be associated with light intensity. One of the environmental signals that activate tuberization in potatoes is light. Delivery of light from the light-receiving organ, the leaf, to an underground organ growing in the dark, the stolon, leads to the formation of potato tuber [27] [36] . However, the underground tubers are exposed to darkness while the photosynthetically active leaf tissues are exposed to the light. This reason, in addition to the reduced DHAR activity in the dark, justify for a reduced DHAR activity in tuber tissues than shoot, and thus reduced amounts of reduced ascorbate in tubers compared to the shoot in this study.
4.4. Loses of Bioactive Compounds during Potato Processing
When different processing steps were compared, significant losses of total phenolics, anthocyanins and total antioxidant capacity took place during the peeling (18% - 23%) and blanching processes (35% - 60%). With further cooking/reconstitution, additional losses ranged between 7% - 12% with no prominent genotypes differences. Together, 65% to 90% of these bioactive compounds were lost during processing. The levels of anothocyanins were found to be affected more in the processing compared to total phenolics and antioxidant capacity. Cultivars with greater initial levels of phenolics (coloured genotypes) lost about 65% and genotypes those had less phenolics to start with (unpigminted genotypes), lost about 50% to blanching.
Significant amounts of bioactive compounds in potato peels also have been reported by others. Ji et al. [37] reported purple and red pigmented potato clones contained higher levels of phenolics and anthocyanins, and had higher antioxidant capacity in both peel and tuber than observed in yellow or unpigmented clones. Samarin et al. [38] also reported significant amounts of phenolic compounds in potato peels, adequate for commercial extraction. Lewis et al. [15] found that purple- and red-skinned tubers contained twice the concentration of phenolic acids as white-skinned tubers. Genotype differences in peeling losses of bioactive compounds were prominent in our study with maximum losses from POROIPG22-1 (phenolics 28%, anthocyanins 25% and antioxidant capacity 32%; Table 1). This was due to the smaller tuber size (98 g/tuber) of this genotype (Table 1). Though all tubers were peeled with a standard kitchen peeler at a similar depth (1 - 2 mm), as the tubers were of different size, smaller tubers had more tissue removed with the peel compared to the rest of the tuber tissues.
In general, cooking leads to reductions in bioactive compounds and antioxidant capacity for most vegetables (however varies with the methods of cooking) as has been seen in our study. Other studies also have shown that different cooking treatments reduce various phenolic compounds in potatoes [19] [20] . Lachman et al. [39] reported that cooking treatments (boiling, baking and microwaving) reduced ascorbic and chlorogenic acid contents, total glycoalkaloids, a-chaconine and a-solanine with the exception of total anthocyanins.
Significant loss of total phenolics, anthocyanins and antioxidant capacity during blanching of the current study can be attributed to peeling, the browning phenomenon during cut, leaching, levels of temperature and duration and pH during blanching, selective decomposition, and/or inter-conversion of the compounds [6] [13] . Peeling and slicing of tuber cause browning and expose greater pulp surface to hot water [17] which may facilitate greater leaching of bioactive compounds in water bath during blanching. Perla et al. [19] reported selective leaching of polyphenols from tubers is one of the factors responsible for the decline of polyphenols, flavonoids and flavonols during boiling. They determined that on average, boiling, microwaving and baking reduced total phenolics to 54%, 46.5% and 46%, respectively. Chlorogenic acid, one of the main polyphenols in potatoes, was completely destroyed during baking at 212˚C in an oven for 45 min, reduced to 35% and 55% respectively after boiling for 30 min in water, or microwaving at 218˚C for 30 min and was not detected in commercially processed potato products [20] . Besides altering quantities of polyphenols, food processing that involves heating has been shown to alter flavonols and these alterations directly affected the antioxidant status of plant tissues [16] . Stability of anthocyanins and phenolics are strongly dependant on the pH value in water media and also the temperature [17] . This and hydrophyllic property of anthocyanin [5] probably justify the maximum loses of anthocyanins during blanching in our study.
Different from literature data that indicate unmodified or an increased amounts of phenolics, anthocyanins and antioxidant capacity after cooking [8] [17] , in our study the contents of these bioactive compounds drastically decreased during processing/ cooking. This disagreement may be related to the fact that in those studies the cooking conditions were similar to household cooking and did not involve blanching step as in the commercial processing conditions. In some studies, the tubers were not peeled; also either the levels of cooking temperature was low, the cooking duration was less compared to commercial processing, or there was a combination of these two. Indeed, physical and/or chemical changes during processing may differ depending upon the methods used to process potatoes.
5. Conclusion
Taken together, these findings suggest that the pigmented potatoes had greater amounts of phenolics, anthocyanins and antioxidant capacity compared to unpigmented genotypes. Total ascorbate contents were not associated to tuber flesh colour. However, the bioactive compounds and antioxidant capacity were significantly reduced by peeling and significantly more blanching, without prominent differences among genotypes. This study suggests that blanching treatments would be one of the most challenging areas to focus on to minimize the adverse effects of processing and to obtain processed/reconstituted products that keep more of their original characteristics including novel bioactive compounds. Peeling process also needs some attention. This work is the first of its kind that focused on the cultivars that the industry recommended, and the processing methods used at the industrial settings. The results are impotent to guide the potato processing industry for their product development and revision of their processing methods.
Acknowledgements
The research was supported by New Brunswick Enabling Agricultural Research and Innovation (EARI) of Growing Forward Program, McCain Foods Ltd., Canada and Agriculture and Agri-Food Canada (AAFC). The authors also would like to thank Michele Elliot for technical assistance.