Dimethylsulfide and Coral Bleaching: Links to Solar Radiation, Low Level Cloud and the Regulation of Seawater Temperatures and Climate in the Great Barrier Reef ()
1. Introduction
Reef-building corals contain high concentrations of dimethylsulfoniopropionate (DMSP) [1] - [6] that can be enzymatically converted to dimethylsulfide (DMS), a trace sulfur gas that oxidizes in the atmosphere to form nss-sulfate aerosol particles and cloud condensation nuclei (CCN) (Figure 1). The growth
![]()
Figure 1. The climate feedback involving production of DMS that can occur from coral reefs and adjacent ocean waters of the Great Barrier Reef. Oceanic phytoplankton and coral reefs produce seawater DMS which is transferred by air-sea transfer to the atmosphere where it oxidises to non-sea sulfate aerosol and methane sulfonate (MSA). The tiny sulfate aerosol particles grow to produce cloud condensation nuclei (CCN) forming low level cumulous and stratocumulous clouds, thus decreasing solar radiation levels, SSTs, and increasing rainfall. [Adapted from Charlson et al. 1987 [7] and Jones and Gabric, 2006 [8] ]. Low tides and convective rainfall can increase the flux of DMS to the reef atmosphere, whilst wind speed over the ocean affects DMS transfer to the ocean atmosphere [9] [10] .
of CCN can lead to the formation of low level cloud (LLC) such as cumulous and stratocumulus clouds, decreasing solar radiation and sea surface temperatures (SST), thus influencing climate as a negative climate feedback [7] [11] . Atmospheric DMS can be oxidized to nss-sulfate aerosol by hydroxyl (OH) and nitrate (NO3) radicals [12] , although O3, HO2, and H2O2 can also be involved [13] . The products of this oxidation produce SO2, methanesulfonate acid (MSA) and gas-phase sulfuric acid (H2SO4) which can then condense onto aerosol particles [14] , or nucleate to form new sulfuric acid particles [15] (Figure 1). The climate impact of atmospheric DMS (DMSa) depends on the ability of its oxidation products to increase CCN, cloud droplet concentration [13] , LLC and rainfall [16] [17] .
The CLAW hypothesis (named after Charlson, Lovelock, Andrea and Warren) [7] states that because the albedo of low level clouds (Figure 1), and thus the Earth’s radiation budget is sensitive to CCN number; biological regulation of the climate is possible through the effects of sea surface temperature (SST) and sunlight on phytoplankton population and DMS production [7] . This climate regulation is also thought to occur over coral reefs [18] [19] [20] . Variations in solar radiation can increase or decrease phytoplankton activity which in turn can increase or decrease seawater and atmospheric DMS [21] . This may increase or decrease the albedo of LLC cover in the marine boundary layer (0 - 1000 m), which will decrease or increase SSTs, thus regulating climate [7] . Charlson et al. [7] hypothesized that as temperatures increase from GHG warming, phytoplankton DMS production will increase, thus increasing CCN and cloud albedo, decreasing solar radiation and lowering SSTs, keeping global temperatures relatively constant; a negative climate feedback (Figure 1). Such a cloud cover-SST feedback occurs in the NE Pacific [22] , and over coral reefs in the GBR [23] , but it is not known whether DMS is implicated in these feedbacks. An increasing amount of evidence now suggests that a LLC climate feedback involving enhanced production of DMSa and other aerosols from coral reefs could occur in the GBR [18] [20] , and indeed over reefs in the western Pacific warm pool (WPWP) [24] [25] . For example, air masses that travel over coral reefs in the southern GBR contain ammonium sulfate aerosols (Figure 1) derived mainly from the oxidation of DMSa which are likely produced from the GBRs coral reefs [26] . Modini et al. [26] have characterised these freshly formed aerosol particles that consist of 60% ammonium sulfate and 40% organic material. These measurements are the first direct observation of aerosol particle formation over the GBR, and compare with earlier observations [27] . Mean aerosol particle concentrations averaged 3200 cm−3 and consisted of broad pulses of aerosol particles emitted in the morning and afternoon, with a small discrete aerosol burst at mid-day [26] , a distribution similar to that measured for DMSa over coral reefs in the central GBR [28] . Periodic pulses of DMSa often occur over coral reefs during very low tides and rising tides [10] [28] [29] [30] . It is apparent that natural stress events (e.g. low tides, high SSTs, rainfall) cause the coral to utilize intracellular DMSP to cope with the resultant osmotic, solar radiation and temperature stresses [31] [32] [33] , and in the process produce enhanced levels of seawater and atmospheric DMS [10] [28] [30] [34] .
These periodic pulses of DMSa at low tides may be capable of increasing LLC cover over coral reefs (Figure 1), keeping SSTs < 30˚C, thus decreasing the incidence of coral bleaching and significantly affecting regional climate in the GBR, western and central Pacific Ocean [20] [23] [24] [28] [35] . However, the increased frequency of coral bleaching in the GBR in the last two decades could suggest that the strength of this feedback is decreasing [36] [37] [38] [39] [40] .
This article discusses the potential of a “coral reef-DMS-cloud” feedback involving reef production of DMSa to constrain SSTs in the GBR, and could be the reason why coral reefs in the WPWP have not bleached so frequently over a period of 25 years [24] . In early January 1994 very high seawater temperatures (>33˚C), coupled with high air temperatures (44.3˚C), associated with hot westerly winds, occurred at Nelly Bay reef, Magnetic Island off Townsville in the central GBR. Many colonies of corals at this location bleached to a pale colour and colonies of some species (mainly Acropora and Pocillopora) bleached to a bone-white colour [31] [41] . In contrast, no coral bleaching was recorded at Pioneer Bay, Orpheus Island 70 km to the north of Townsville, reflecting the lower SSTs at this study site [28] . Prior, during and after the severe bleaching event at Nelly Bay seawater DMS and DMS fluxes were measured at both Pioneer Bay and Nelly Bay fringing coral reefs. The DMS database gathered during these bleaching events are unique in that it is the only DMS database we have before, during and after a coral bleaching event in the GBR. Given that DMS could have such an important role in the development of LLC cover over coral reefs, thus regulating SSTs, the aims of this paper are: 1) To highlight the changes in dissolved or seawater DMSw, dissolved NH4+ and DMS flux, solar radiation and SST before, during and after the bleaching event. 2) To examine the changes in LLC cover with low tides and rainfall before, during and after the bleaching event. 3) To examine any correlations or links between LLC cover, solar radiation, SST, rainfall and the sulfur substances measured in order to throw more light on this possible coral reef climate feedback in the GBR. Whilst further studies are necessary on the physics and chemistry of sulfate aerosol, CCN and LLC formation over coral reefs in the GBR, the study described here provides more momentum for such studies to take place and may provide more insights into the frequency and intensity of this coral reef-climate feedback, as well as it’s sustainability in the face of GHG warming and increased coral bleaching.
2. Study Areas and Coral Bleaching
2.1. Nelly Bay-Magnetic Island
Nelly Bay is one of the larger embayments on the eastern coast of Magnetic Island approximately 7 km from Townsville and faces SE into Cleveland Bay (Figure 2). The approximate size of the reef at Nelly Bay is 43 hectares. Within Nelly Bay the reef rises sharply from the general Cleveland Bay floor to form a wide
![]()
Figure 2. Map of the study area showing locations of Nelly Bay fringing reef on the NE coast of Magnetic Island and the Pioneer Bay reef site at Orpheus Island north of Townsville.
reef flat that dries at lowest low tides (Figure 2). The reef slope area contains areas of abundant coral cover mixed with other areas dominated by brown algae. The study site (146˚51'N, 19˚10.5'E) was approximately 700 m directly offshore. This site was chosen since it could be easily accessed from the shore, and contained many species of corals that contained high concentrations of DMSP, and so produced high concentrations of DMSw compared with phytoplankton production [3] . Human impacts such as increased suspended sediments from dredging and enhanced nutrient levels from sewage discharge into Cleveland Bay affect this site [42] [43] .
2.2. Pioneer Bay-Orpheus Island
Orpheus Island (lat. 18˚35'S, long. 146˚29'E) is ~70 km NE of Townsville on the east coast of Queensland (Figure 2). A major seasonal study was conducted at this site over 19 months, with 18 regular visits to the James Cook University marine research station at Pioneer Bay Reef between 7 December 1992 and 25 February 1994, and one further visit on 19 July 1994 [28] . The DMS results of this study are summarised here in order to make comparisons with Nelly Bay reef, and because the two reefs together give a more accurate idea of the flux of DMS from coral reefs in the local area during the study period. The sampling location for the present study was at the edge of the fringing coral reef in the northern part of Pioneer Bay. The fringing reef flat is ~400 m wide and the sampling site was specifically chosen because both the flooding and ebbing tide travel parallel with the coast over a significant amount of reef before reaching the sampling point [44] . At this site, there was enough water above the reef that even at the lowest of tides, the reef was just covered, enabling water sampling at all tidal heights. The reef flat and slope at Pioneer Bay contain a diverse assemblage of corals, although the coral community is likely impoverished because of human impacts from the marine station.
2.3. Coral Bleaching
The 1994 coral bleaching event at Nelly Bay, Magnetic Island was first observed by the Great Barrier Reef Marine Park Authority (GBRMPA) on 16th January [44] . Underwater observations of the 1994 bleaching event at Nelly Bay from 28th January to 14th February have been reported [44] [45] . Jones (1995) [44] observed that the most pronounced bleaching occurred at this site in the arborescent and tabulate acroporids, the montiporids and the pocilloporids from 8th January. In some cases heavily bleached colonies were observed immediately adjacent to normal coloured colonies of the same species. In the heavily bleached branching coral species, there were usually slight tinges of yellow/brown colour on the undersides of the branches, especially those in the interior of the colonies. Preferential loss of colour on the upward facing surfaces of the corals repre- sented a frequent bleaching response and indicated an effect from increased levels of solar radiation [46] . The first signs of recovery from the bleaching event were noted on a dive conducted on 24th February 1994 [44] . Mottled or paler patches on partially bleached colonies appeared slightly browner and colonies readily regained their colouration in the 3 - 4 months after the bleaching event. By May 1994 few discoloured colonies could be observed at Nelly Bay, although pale yellow colonies of Pocillopora damicornis were observed as late in the year as July 1994 [31] [45] . Many colonies of P. damicornis lost in excess of 90% of their zooxanthellae, and bleached to a bone white colour. Six months after the bleaching event many colonies of P. damicornis were still bleached with zooxanthellae densities up to 10 times lower than unbleached colonies (1.22 × 105 compared to 2.67 × 106 cm−2). Changes in intracellular DMSP concentrations in this coral have been recently reported [31] . In contrast to Nelly Bay no bleaching of corals at Pioneer Bay was observed during our study period.
3. Methods
3.1. Sea and Air Temperatures, Solar Radiation, Cloud Cover, Rainfall and Wind Speed
Continuous sea surface temperatures (SST) were measured at Geoffrey Bay (next bay to Nelly Bay), Magnetic Island by staff from AIMS which enabled heating and cooling periods to be expressed as degree heating week (DHW) or degree cooling weeks (DCW) over the November to March period. Average daily air temperatures, cloud cover and rainfall were recorded by the Townsville Bureau of Meteorology (TBM). Daily global solar radiation and low level cloud (LLC) cover was measured at Townsville airport, about 5 km from Nelly Bay by the Bureau of Meteorology (BoM) and downloaded from the BoM website. LLC was the amount of the sky covered by cloud present in the marine boundary layer (MBL = upper 500 - 1000 m) and is an estimate of marine cumulous and stratocumulus cloud which DMSa is reputed to form [7] . LLC measurements were made every 3 hours and averaged to give daily mean values. The daily global solar exposure is the total solar energy for a day falling on a horizontal surface. It is measured from midnight to midnight. The BoM’s computer radiation model uses visible images from geostationary meteorological satellites to estimate daily global solar exposures at ground level. At each location the image brightness is used to provide an estimate of the solar irradiance at the ground or sea surface. Essentially, the irradiance at the surface can be calculated from the irradiance at the top of the earth’s atmosphere, the amount absorbed in the atmosphere (dependent on the amount of water vapour present), the amount reflected from the surface (surface albedo) and the amount reflected from clouds (cloud albedo). These instantaneous irradiance values are integrated over the day to give daily solar exposure in MJ・m−2 and are clearly a reflection of cloud cover, aerosol concentrations and water vapour in the atmosphere. Both LLC and solar radiation measurements whilst measured at Townsville airport are assumed to reflect conditions in the local geographical area.
3.2. DMSw,  and DMS Flux Measurements
and DMS Flux Measurements
Water samples from Nelly Bay, Magnetic Island were collected via a small boat about 700 m offshore and over the coral reef. The site was marked by a small buoy [43] . Seawater was gently filtered using a 50 mL silanized glass syringe, with Teflon plunger, through a pre-rinsed 0.45 µm cellulose acetate filter (Minisart, Sartorius). Water samples from Nelly Bay and Pioneer Bay were collected and analysed for DMSw according to procedures already described [28] [46] . The flux of DMS from Nelly Bay and Pioneer Bay fringing reefs was calculated using the method of Liss and Merlivat (1986) [9] and calculated as reported in Curran and Jones (2000) [47] . More continuous measurements of DMSw and DMS flux at Nelly Bay reef were obtained by using the regression equations between DMSw and SSTs (DMSw = 0.5539 SST − 12.074; r2 = 0.44, p < 0.001), and DMSw and DMS flux (DMS flux = 2.247 DMSw − 1.6945; r2 = 0.67, p < 0.01), enabling comparison with actual measurements. Calculated DMSw and DMS flux values compared very favorably with values obtained from seawater DMS measurements and application of the transfer velocity equation [9] (in prepn.). Samples collected from Nelly Bay were analyzed for 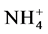 by AIMS [43] .
by AIMS [43] .
3.3. Zooxanthellae Densities in Acropora formosa Coral
Zooxanthellae densities in A. formosa have been reported [44] . They are replotted here as a proxy of the stresses that affected corals at Nelly Bay and provide some insights on the influence of A. formosa on DMS in seawater given that coral zooxanthellae produce large amounts of DMS [20] . Zooxanthellae densities in Pocillopera damicornis, which was still bleached at Nelly Bay in June 1994, have recently been reported, as well as intracellular levels of DMSP [3] [31] .
3.4. Statistical Analysis
SPSS 11 was used for the statistical analysis of the data sets. For the seawater data, statistically significant differences between treatments were tested using a Pearson’s Product Linear Correlation analysis (parametric, normally distributed) with significance determined at the 0.01 to 0.001 level.
4. Results
4.1. Solar Radiation and Air Temperatures
Periods of very high solar radiation levels occurred over 29 days (>29 MJ・m−2) in November (DOY 1-31), December (DOY 31-61) and early January (DOY 62- 72), with 9 consecutive days of extreme solar radiation levels from 27th November to 5th December (DOY 27-36) when values ranged from 29.2 to 30.9 MJ・m−2・d−1 (mean 30.3) (Figure 3(a)). From 1st to 6th January (DOY 62-71) prior to the bleaching event on 8th January (DOY 69), extreme solar radiation levels ranged from 28.3 to 29.9 MJ・m−2・d−1 (mean 29.4). The mean solar radiation levels over Nelly Bay reef in January (mean = 26.7 MJ・m−2・d−1), with the early January and late November to early December values, were higher than the mean January levels for the 1990-2012 period (24.7 MJ・m−2・d−1), and higher than the highest monthly mean for all 22 years (28.4 MJ・m−2・d−1) (BoM online data). From 4-9th January (DOY 65-70), daily air temperatures averaged over 30˚C, and included a series of exceptionally high maximum daytime temperatures in excess of 36˚C recorded by the Townsville Bureau of Meteorology (TBM). On 7th January 1994, the maximum air temperature was 44.3˚C, the highest air temperature recorded at the time by the TBM since 1942 [44] . From 19th-23rd January
![]()
![]()
![]()
Figure 3. (a) Solar radiation levels in MJ・m−2・d−1, (b) Mean daily low level cloud cover (oktas), versus day of year (DOY), (c) SST (˚C), lowest nightime tides (m), LLC (oktas) and rainfall (mm) recorded from November to April by the BoM at Townsville airport and Nelly Bay.
(DOY 80-84), daily air temperatures also averaged over 30˚C, and included a series of unusually high minimum daily temperatures, including the second highest minimum air temperature (29˚C) recorded locally since 1942. Daily air temperatures then fell sharply from 29.3˚C on 24th January, to 24.8˚C on 31st January 1994 [44] .
4.2. Low Level Clouds, SST and Rainfall
Regular pulses of LLC cover occurred in November and December that lasted for about a week (Figure 3(b)). From November-December mean daily LLC generally averaged 2 - 3 oktas with pulses often coinciding with low tides and rainfall (Figure 3(c)). These pulses of LLC coincided with relatively stable SSTs of 27.5˚C - 29.5˚C (mean 28.37˚C), except from 19-28th December when SSTs reached 30.3˚C. From the 2nd to 9th January (DOY 63-70) mean daily LLC cover markedly decreased (~1 okta) and coincided with the extreme solar radiation levels that caused corals to bleach white on their upper surfaces at Nelly Bay reef [44] with SSTs increasing to 32˚C - 34˚C which continued to 23rd January (Figure 3(c)). From 10th January to 26th February (DOY 71-118) increasing pulses of LLC cover occurred (2 - 6 oktas) as the corals slowly recovered, and then remained low and relatively constant (daily mean ~2 - 3 oktas) up to 9th March (DOY 118-129). As LLC increased in February, and low tides increased in height from February-March, SSTs gradually decreased (Figure 3(c)). Stable SSTs often coincided with significant rainfall that occurred in November (monthly mean 102 mm) (Figure 3(c)), with lower, but more regular, and smaller amounts falling in December (monthly mean 44 mm), which often fell during low tides. On the 10th and 12th January 54 and 12 mm of rain occurred, with 70 mm falling on 31st January, yielding a monthly mean rainfall of 149 mm. In February very high rainfall occurred on 1st and 2nd February (66, 25 mm), with smaller amounts occurring throughout the month, yielding a monthly mean of 212 mm. From 9-13th March (DOY 129-133) mean daily LLC cover rapidly decreased to zero and then rapidly increased to ~ 4.5 oktas over 9 days to 22nd March (DOY 142) when the mean daily LLC cover decreased gradually to about 1.5 oktas on 31st March (DOY 151) (Figure 3(b)). No rainfall occurred in March, and in April only 0.6 mm of rainfall was recorded on 28th April.
4.3. Variation in DMSw, DMS Flux,  and Coral Zooxanthellae
and Coral Zooxanthellae
DMSw concentrations at Nelly Bay Reef ranged from nd-3.9 nM (mean 2.2 nM) with generally elevated concentrations (>1 µM) occurring from 6th November to the 3rd February, decreasing to very low concentrations (<1 µM) from 13th February to 2nd May (Figure 4(a)). During mid-February to early May DMSw concentrations were particularly low at Nelly Bay (nd-0.6 nM, mean = 0.2 nM) (Figure 4(a)). DMS flux concentrations ranged from nd-4.8 µmol・m−2・d−1 (mean 1.64) with high values before the bleaching event (Figure 4(a)), and elevated flux levels after the bleaching event on 21st January, decreasing to nd-0.8 µmol・m−2・d−1 from 3rd February to 2nd May. A measure of DMS emitted from coral reefs in the wider local region was obtained by combining the DMSw and DMS flux measurements from the fringing coral reef at Nelly Bay fringing reef (Magnetic Island) with those of Pioneer Bay (Orpheus Island) (Figure 4(b)). These measurements again highlight increasing pulses of DMSw and DMS flux that occurred at these two reefs over the period November to the end of February. For example from 4th November-17th November DMS flux emitted from these two coral reefs ranged from 0.4 - 4.4 µmol・m−2・d−1 (mean = 2.4). From 8th January to 3rd February much more DMS was emitted from Pioneer Bay reef than Nelly Bay reef with fluxes ranging from 0.4 - 14 µmol・m−2・d−1 (mean 4.1). On the 24th January the huge pulse of DMSa emitted from Pioneer Bay reef (14 µmol・m−2・d−1) occurred when significant rainfall occurred in the region (10 mm at Townsville airport on 23rd January) and low tides occurred at night (0.9 m) (Figure 3(c)).
Similarly on 25th February an even higher flux of DMSa was emitted from the fringing reef at Pioneer Bay ranging from 19 - 20 µmol・m−2・d−1 (mean 19.5), when medium low tides occurred (0.8 m) (Figure 4(b)). These huge fluxes of DMS emitted from Pioneer Bay reef were in contrast to the very low fluxes of DMS that were emitted from 3rd February to 2nd May from the Nelly Bay fringing reef (nd-0.8 µmol・m−2・d−1) where coral bleaching occurred (Figure 4(a)), and calculated fluxes from 10th January to 23rd January (Figure 4(d)). As the lowest nighttime tides decreased at both Nelly Bay and Pioneer Bay reefs seawater
DMSw increased (Figure 4(c)), suggesting that DMS fluxes increase from reefs as more of the reefs are exposed at low tides. Ammonium concentrations in Nelly Bay from 6th November 1993 to 2nd May 1994 ranged from 0.03 - 1.83 µM (Figure 4(e)), with elevated levels in early November, decreasing to very low levels in early to mid-December, and then a huge increase from the 8th January (1.83 µM) during the coral bleaching period. Very high concentrations (>1 µM) occurred up to 31st March, with levels decreasing again to very low levels in April and May (Figure 4(e)). Zooxanthellae densities in Acropora formosa started to rapidly decrease on 8th January as this coral started to bleach, reaching lowest levels on 13th February [44] (Figure 4(f)). On 31st March as SSTs decreased to 27˚C at Nelly Bay, zooxanthellae densities reached 6.5 × 104 zooxanthellae per polyp, and then slowly increased their numbers, stabilizing at 10.5 × 104 zooxanthellae per polyp on 2nd May (Figure 4(f)). These concentrations are still very low for this coral suggesting this coral had not completely recovered from the bleaching, and may even have been bleaching in November to December. At Pioneer Bay, Orpheus Island no bleaching of corals was reported.
4.4. Continuous SST Record
A continuous record of SSTs was recorded by AIMS at Geoffrey Bay (Magnetic Island) at 2.1 m depth on the reef flat. Several warming, cooling and stable periods of SSTs were evident over the November to March period (Figure 5). For example, from 1st-9th November SSTs increased from 27.55˚C - 28.81˚C at a rate of 0.11˚C per day. SSTs then remained relatively stable from 9th-17th November
![]()
Figure 5. SST (˚C) for Magnetic Island reef flat seawater from November-April. Heating and cooling periods, together with stable SST periods are highlighted, with the thickness of the arrows indicating the magnitude of the cooling and warming.
(mean 28.61˚C ± 0.16˚C) until another warming period occurred from 17th-22nd November with a heating rate of 0.17˚C per day. From 22nd-25th November another stable SST period occurred (mean 29.49˚C ± 0.11˚C), followed by a cooling period from 25th November to 1st December when SSTs cooled from 29.52˚C to 27.31˚C at a rate of 0.28˚C per day. This was followed by yet another period of stable SSTs from 2nd-10th December (Mean 28.11˚C ± 0.38˚C). From 10th-14th December another cooling period occurred with SSTs decreasing from 28.32˚C to 27.2˚C at a cooling rate of 0.22˚C per day. From 15th-19th December SSTs were relatively stable at 27.54˚C ± 0.25˚C. From the 18th December to 27th December, a period of 10 days, SSTs increased from 27.37˚C to 30.32˚C (Figure 5) increasing at a rate of ~0.30˚C per day. From 28th to 31st December a stable period of SSTs again occurred ranging from 28.02˚C - 29.40˚C (mean = 28.71˚C ± 0.6˚C). SSTs from 2nd to 9th January rapidly increased from 30.3˚C to 34˚C (mean 31.7˚C) at a rate of ~0.46˚C per day. From 10th-15th January SSTs cooled significantly (0.38˚C per day) to 30.71˚C on 15th January. Another stable but high SST period then occurred from 15th to 21st January, with SSTs stabilizing at 31.87˚C ± 0.25˚C (above the coral bleaching threshold), followed by a short, sharp warming period from 21st-23rd January when SSTs reached 31.95˚C (increasing at 0.22˚C per day). A sustained and rapid cooling period then occurred from 23rd January to 3rd February, decreasing to 27.75˚C at a rate of 0.20˚C per day (Figure 5). From 3rd-6th February SSTs rapidly increased at the highest warming rate of 0.39˚C per day, peaking at 29.56˚C on 6th February. This was followed by the longest stable period of SST s from 6th February to 23rd February (mean 29.30˚C ± 0.28˚C) when more regular pulses of LLC occurred (DOY 98-125; Figure 3(b)), and when significant amounts of rain fell over this period (Figure 3(d)). From 24th February to 12th March a second warming period occurred at a much lower warming rate of 0.1˚C per day (28.72˚C - 30.46˚C), followed by decreasing SSTs from 12th March (30.26˚C) to 29th March (25.81˚C). SSTs then cooled to 24.9˚C on 4th May.
4.5. Low Level Clouds and Links to Low Tides and Rainfall
Mean daily LLC cover over Cleveland Bay reefs was compared with changes in the lowest nighttime and daytime tides (Figures 6(a)-(e)), and presence of rainfall (Figure 3(d)), as these natural factors can release large amounts of DMSa from coral reefs [10] [29] . These large pulses of DMSa can potentially assist LLC formation (see Figure 1). Increasing pulses of LLC occurred from 1-25th November and often coincided with very low nighttime tides (−0.1 to 0.8 m) (Figure 6(a)), with 25 mm rain falling between 13th-17th November during some of the lowest nighttime tides of the month (−0.1 - 0.5 m) (Figure 3(c)). Pulses of LLC cover also occurred from 16th-21st November on rising tides of 0.2 m to 1 m (no rainfall), whilst from 23rd-26th November a large pulse of LLC occurred during low nighttime tides of 0.5 - 0.6 m and high rainfall (77 mm) (Figure 6(a), Figure 3(c)). On 30th November high levels of LLC coincided with a low nighttime tide of 0.5 m and 4 mm of rainfall. The lowest daytime tides from November-February were always higher than the lowest nighttime tides with both low tides often coinciding with elevated LLC (Figures 6(a)-(e)). In December, tides were again very low, mostly at night. Pulses of LLC cover often coincided with decreasing lowest nighttime tides (Figure 6(b)) and very high DMS fluxes of 6 - 8.6 µmol・m−2・d−1 (Figure 4(d)), in the more continuous flux profile, and more regular rainfall (Figure 3(c)). Rainfall was not as high as in November but occurred intermittently throughout December and often occurred during periods of the lowest nighttime tides (−0.1 to 0.6 m; Figure 3(c)). In the first week of January when corals bleached at Nelly Bay there was almost no LLC cover in the Townsville region (Figure 6(c)), whilst DMS fluxes were low ranging from 0.4 - 3.4 µmol・m−2・d−1 (Figure 4(a)) during medium low nighttime tides of ~1 m. From 9th January increasing pulses of LLC cover occurred over coral reefs in the Townsville region and again often occurred during periods of the lowest nighttime tides (0.3 and 0.4 m) and intense bursts of tropical rainfall on 10th January (54.4 mm) and 12th January (9.8 mm) (Figure 3(c)). From 22nd to 30th January rainfall ranged from 0.2 mm to 10.4 mm with the highest rainfall occurring on a 1m low tide on 23rd January (Figure 3(c)). On 31st January to 2nd February the highest levels of LLC cover occurred (3 - 5 oktas) during intense bursts of tropical rainfall and coincided with relatively high nighttime low tides (1 - 1.3 m) (Figure 6(c), Figure 6(d), Figure 3(c)). More regular pulses of LLC cover
![]()
Figure 6. Mean low level cloud (LLC) in oktas and lowest daytime and nighttime tides (m) for the Townsville-Cleveland Bay region from November to March (a)-(e).
often occurred during low tides after 2nd February, and often coincided with rainfall bursts of <10 mm (Figure 6(d)). Smaller differences in low tides occurred between the lowest nighttime tides and lowest daytime tides in February (Figure 6(d)) and these prolonged periods of low tides and heavy rainfall (>70 mm) may have caused the extremely large DMS fluxes (20 µmol・m−2・d−1) from Pioneer Bay on 25th February (Figure 6(d), Figure 4(b), Figure 3(c)). In March the lowest nighttime tides ranged from 0.6 - 0.8 m with pulses of LLC cover often coinciding with lowest tides from 1st-6th March (Figure 6(e)), whilst high LLC cover occurred in the third week of March during decreasing daytime and nighttime low tides (1.5 m - 0.5 m) (e.g. 18th-24th March). No rainfall occurred in March.
4.6. Alternating Warming and Cooling Periods
The heating and cooling periods of the Nelly Bay reef flat over the 5 month period (Figure 5) were first calculated as changes in SST per day and then were scaled up to “Degree Heating Week” (DHW) or “Degree Cooling Week” (DCW) as coral ecologists often use a similar index to compare coral bleaching events around the world. We have done this so that we can compare each warming and cooling period over a standard amount of time (i.e. 7 days). For example, the first heating period from 1-9th November (DHW +0.77˚C) was followed by relatively stable SSTs from 9-17th November (28.61˚C ± 0.16˚C) (Figure 7). This was followed by a heating period from 17-22nd November with a DHW of +1.19˚C followed by a SST stabilization period from 22-25th November of 29.49˚C ± 0.10˚C, followed by a significant cooling period from 25th November to 1st December with a DCW of −1.96˚C which surprisingly was equal to the sum of the two previous DHWs (i.e. +0.77 +1.19 = +1.96˚C). These SST heating and cooling periods in November (Figure 7) were generally accompanied by elevated levels of DMSw, DMS flux at Nelly Bay and Pioneer Bay (Figure 4(a), Figure 4(b)), elevated LLC (Figure 6(a)) and intermittent rainfall (Figure 3(c)), during periods of low daytime and nighttime tides (0.26 - 0.83 m, mean 0.59 m) (Figure 6(a)). Over 9 days from 2-10th December (Figure 7) another SST stabilization period occurred (28.11˚C ± 0.38˚C). From 10-14th December another cooling period occurred with a DCW of −1.54˚C, which was again followed by a SST stabilization period from 15-19th December (27.54˚C ± 0.25˚C) (Figure 7). From 19-28th December the second highest warming event occurred (Figure 5) giving a DHW of +2.31˚C, with SSTs at 30.3˚C, just over the thermal bleaching threshold of Acropora. This was followed by a SST stabilization period from 28-31st December when SSTs were ~28.7˚C. From 15th-31st December DMSw and DMS flux gradually increased at Nelly Bay reef (Figure 4(d)) in the more continuous record, whilst LLC increased from 17-23rd and 28-31st December (Figure 6(b)). These periods of enhanced DMSw and DMS flux in December often occurred during the lowest nighttime tides (0.1 - 0.7 m; mean 0.52 m) and periods of high LLC (Figure 6(b)). From 2-9th January mean SSTs reached 31.7˚C with a DHW of +1.4˚C, followed by a short rapid cooling period from 10-15th January of
![]()
Figure 7. DCW (white) and DHW (red) in ˚C week−1 and stable SSTs (green) (˚C) for Magnetic Island reef flat waters at 2.1 m depth for the summer period November to March.
−2.66˚C (Figure 7). This was followed by high mean SSTs from 15-21st January of 31.9˚C and relatively high DMSw and DMS fluxes from the reefs (Figure 4(a), Figure 4(b), Figure 4(d)).
From 21-23rd January a short warming period occurred with a DHW of +1.54˚C coinciding with enhanced DMS fluxes from Pioneer Bay reef (14 µmol・m−2・d−1) (Figure 4(b)), and increasing levels of LLC from 21st January to 3rd February (Figure 6(c), Figure 6(d)). This was followed by a similar magnitude cooling period from 23rd January to 3rd February of −1.4˚C (Figure 7). A short intense warming period from 3-6th February recorded the highest DHW of +2.73˚C when DMSw and DMS fluxes were very low on 3rd February (0.7 µmol・m−2・d−1) (Figure 4(a)) and corals were still stressed as evidenced by low zooxanthellae counts in Acropora (Figure 4(f)). From 6th-23rd February an exceptionally long period of stable SSTs of 29.3˚C ± 0.28˚C occurred (Figure 7), coinciding with regular pulses of LLC (Figure 6(d)) and high DMS fluxes at Pioneer Bay reef (14 µmol・m−2・d−1) in late January, and on 25th February) (20 µmol・m−2・d−1) (Figure 4(b)), and in the more continuous flux record (Figure 4(d)). The Pioneer Bay reef did not experience bleaching since SSTs were < 30˚C over the 19 month study, whilst at Nelly Bay reef in early to mid-February DMS flux was very low at ~0.4 µmol・m−2・d−1 and SSTs were exceptionally high 30˚C - 34˚C. A small warming period of +0.7˚C DHW occurred from 24 February-12th March, followed by a cooling period of −0.42˚C as the region exited the warm summer months.
4.7. Regression Analysis
Solar radiation was significantly positively correlated with DMSw and DMS flux (p < 0.05) indicating that as solar radiation increases both DMSw and DMS flux also increase (see Table 1). One of the most interesting findings in this work was
![]()
Table 1. Regression analysis and level of significance for Nelly Bay and Pioneer Bay.
Units: DMSw = seawater DMS (nM). SR = Solar radiation (MJ・m−2). DMS flux = µmol・m−2・d−1. ![]() = µmol. Zooxanthellae concentration = zooxanthellae polyp−1 × 104. LLC = Low Level Cloud (oktas). LNT = Lowest night-time tide (m). NB = Nelly Bay. PB = Pioneer Bay. * = highly significant. # = Highest DMS flux values for PB omitted in correlation. ## = SST < 30˚C.
= µmol. Zooxanthellae concentration = zooxanthellae polyp−1 × 104. LLC = Low Level Cloud (oktas). LNT = Lowest night-time tide (m). NB = Nelly Bay. PB = Pioneer Bay. * = highly significant. # = Highest DMS flux values for PB omitted in correlation. ## = SST < 30˚C.
the very significant positive correlation of DMSw with SST (r2 = 0.44, p < 0.001) for Pioneer Bay fringing reef (see Table 1). However, there was no significant correlation of DMSw and SSTs for Nelly Bay reef if we included SSTs > 30˚C. However, when SSTs > 30˚C are excluded there was a significant positive correlation between DMSw and SSTs for Nelly Bay reef (r2 = 0.83, p < 0.01). Interestingly almost the same linear regression equation was obtained for the DMSw- solar radiation correlation for Nelly Bay reef as the DMSw-SST correlation for Pioneer Bay reef (see Table 1). However, the DMSw-SST linear regression equation for Nelly Bay had a much greater slope by as much as 162%, compared with Pioneer Bay. It is interesting that the regression equation that correlates the lowest nighttime tide with DCW and DHW (Figure 8(a)), although just short of significance at p > 0.1, indicates that cooling of reef flat waters generally occurs during low nighttime tides that ranged from 0.5 - 0.9 m (mean 0.5 m). In contrast warming of reef flat waters generally occurred during tides that ranged from 0.7 - 1.4 m (mean 0.97 m) which seems the reverse of what you might expect. Also the regression equation that describes the significant (p < 0.10) relationship between DMS flux and DCW and DHW (Figure 8(b)) indicates that cooling of reef flat waters generally occurred at lower DMS fluxes that ranged from 0.1 - 7.48 µmol・m−2・d−1 (mean 4.8), whilst warming of reef flat waters generally occurred at slightly higher DMS fluxes that ranged from 4.4 - 7.8 µmol・m−2・d−1 (mean 6.4). DMS flux decreased linearly with DCW, although the correlation was not significant at p > 0.10 (Figure 8(b)). DMS flux was very significantly (p < 0.001) correlated with reef flat SSTs (r2 = 0.80) for SSTs ranging
![]()
Figure 8. Results for Nelly Bay fringing reef flat. (a) DHW and DCW plotted against the lowest nighttime tides (m). (b) DMS flux (µmol・m−2・d−1) versus DCW/DHW (˚C week−1). (c) DMS flux (µmol・m−2・d−1) versus reef flat SSTs (˚C). (d) LLC versus DMS flux (µmol・m−2・d−1).
from 27.5˚C to 33.3˚C, with the correlation described by a polynomial equation (n = 2) (Figure 8(c)). As SSTs breached the coral bleaching threshold and reached maximum SSTs of 33.3˚C, DMS flux decreased to nd levels (Figure 8(c)). When SSTs > 30˚C were omitted from the correlation DMS flux was significantly (p < 0.01) linearly correlated with increases in SSTs from 27.5˚C to 29.68˚C (Figure 8(c)). Seawater ![]() was also significantly positively correlated with SSTs at Nelly Bay reef (r2 = 0.56, p < 0.05) (Table 1). DMSw measured at Nelly Bay and Pioneer Bay reefs was significantly positively correlated with the lowest nighttime tides (r2 = 0.57, p < 0.02) with the correlation predicting that as the lowest nighttime tide decreases, DMSw often increases. Also of interest was the highly significant positive correlation of DMSw and DMS flux for Nelly Bay and Pioneer Bay (r2 = 0.67, p < 0.001) (see Table 1). Whilst the correlation between zooxanthellae counts in Acropora formosa and DMS flux was not quite significant (p > 0.10), more data points may establish a statistically significant correlation. However, the correlation suggests that as zooxanthellae counts decrease in Acropora coral the flux of DMS from reefs decreases. There was also a significant correlation of
was also significantly positively correlated with SSTs at Nelly Bay reef (r2 = 0.56, p < 0.05) (Table 1). DMSw measured at Nelly Bay and Pioneer Bay reefs was significantly positively correlated with the lowest nighttime tides (r2 = 0.57, p < 0.02) with the correlation predicting that as the lowest nighttime tide decreases, DMSw often increases. Also of interest was the highly significant positive correlation of DMSw and DMS flux for Nelly Bay and Pioneer Bay (r2 = 0.67, p < 0.001) (see Table 1). Whilst the correlation between zooxanthellae counts in Acropora formosa and DMS flux was not quite significant (p > 0.10), more data points may establish a statistically significant correlation. However, the correlation suggests that as zooxanthellae counts decrease in Acropora coral the flux of DMS from reefs decreases. There was also a significant correlation of ![]() with zooxanthellae counts (r2 = 0.50, p < 0.10), and an almost significant correlation between DMSw and zooxanthellae counts (r2 = 0.43, p > 0.10). Whilst the overall correlation between
with zooxanthellae counts (r2 = 0.50, p < 0.10), and an almost significant correlation between DMSw and zooxanthellae counts (r2 = 0.43, p > 0.10). Whilst the overall correlation between ![]() and DMSw was not significant, when data points from the 13th February are not included (i.e. when Acropora reached it’s lowest zooxanthellae concentrations) the correlation was highly statistically significant (r2 = 0.99, p < 0.001). Whilst more data points are needed the DMSw-LLC correlation is highly significant (p < 0.001) and suggests that as DMSw in reef environments increases LLC cover also increases (see Table 1). The correlation between DMS flux and LLC is also highly significant (Figure 8(d)) (see Table 1).
and DMSw was not significant, when data points from the 13th February are not included (i.e. when Acropora reached it’s lowest zooxanthellae concentrations) the correlation was highly statistically significant (r2 = 0.99, p < 0.001). Whilst more data points are needed the DMSw-LLC correlation is highly significant (p < 0.001) and suggests that as DMSw in reef environments increases LLC cover also increases (see Table 1). The correlation between DMS flux and LLC is also highly significant (Figure 8(d)) (see Table 1).
Different LLC-SST correlations occurred for different cooling and heating periods experienced at the Nelly Bay fringing reef. For example, for cooling periods not affected by coral bleaching (i.e. cooling periods 1, 2, 5), increasing levels of LLC from ~0.5 - 4.5 oktas were positively correlated with SSTs (p < 0.001) when SSTs were < 30˚C (Figure 9(a)). However, for cooling periods after coral bleaching (i.e. cooling periods 3, 4), SSTs were negatively correlated with LLC (p < 0.01) and indicated that SSTs could be extremely high (>30˚C) at very low levels of LLC cover (i.e. 2 - 3 oktas) (Figure 9(b)). The regression equation also indicated that this cooling took some time to reduce SSTs to < 30˚C. For the heating periods (i.e. periods 1, 2, 3, 5, 6, 7) that did not include the coral bleaching period 4 (i.e. 2-9th January), increasing LLC was significantly positively correlated with increased SSTs up to 33.5˚C (p < 0.01) (Figure 9(c)). For heating periods that included the coral bleaching event (i.e. periods 1, 2, 3, 4, 7), but not period 5 (21-23rd January) and period 6 (the highest heating rate from the 3rd-6th February), decreasing levels of LLC were accompanied by increases in SSTs as you might expect (Figure 9(d)). Solar radiation levels were significantly correlated with LLC (p < 0.001) (Figure 10). However, although increasing LLC generally attenuated solar radiation there were clearly occasions when extreme
![]()
Figure 9. LLC (oktas) plotted against SSTs for different cooling (a, b) and heating periods (c, d) for Nelly Bay reef flat waters.
solar radiation (~29 - 30 MJ・m−2) occurred at both low levels of LLC (<2 oktas) and medium levels of LLC (3 - 4 oktas) (circled area, Figure 10).
5. Discussion
A detailed comparison of mean daily LLC with the lowest daytime and nighttime tides indicated that on many occasions high levels of LLC often occurred on days
![]()
Figure 10. Mean daily solar radiation levels in MJ・m−2・d−1 are plotted against LLC cover at Townsville airport for the November-April period. [Circled area = values of extreme solar radiation at low and medium LLC].
with the lowest daytime and nighttime tides from November-February (Figures 6(a)-(d)), and often coincided with high sporadic pulses of DMSw and DMS fluxes (Figures 4(a)-(d)). For example, from November-December high levels of LLC often occurred during low tides and high DMS flux (Figures 4(a)-(d), Figures 6(a)-(d)). However, in the first week of January when corals were bleaching at Nelly Bay low levels of LLC occurred (<2 okta) when DMS fluxes from Pioneer Bay and Nelly Bay reefs were very low (0.4 - 1.4 µmol・m−2・d−1 (Figure 4(b) and Figure 4(c)), coinciding with high solar radiation levels and high SSTs (Figure 3(b), Figure 5). From 9th January to 3rd February the flux of DMS emitted from the two reefs ranged from 0.2 - 14 µmol・m−2・d−1 (mean 5), with the highest flux emitted from the more pristine Pioneer Bay reef on 24th January (14 µmol・m−2・d−1) after significant rainfall in the region (10 mm at Townsville airport on 23rd January), and when a nighttime low tide of 0.9 m occurred (Figure 6(c)).
From late January to early December mean daily LLC increased dramatically reaching levels as high as 5 - 6 oktas (Figure 6(d)). Regular pulses of high LLC then occurred for the rest of February, often occurring during low tides and sporadic pulses of very high DMS flux at the Pioneer Bay reef (Figure 4(b)). For example, on 25th February extremely high fluxes of DMSa were emitted from the fringing reef at Pioneer Bay ranging from 19 - 20 µmol・m−2・d−1 (mean 19.5), when medium low tides again occurred (0.8 m) (Figure 4(b)). These huge fluxes of DMS emitted sporadically from Pioneer Bay reef were in contrast to the very low fluxes of DMS that were emitted from 13th February to 2nd May from the Nelly Bay fringing reef (nd-0.8 µmol・m−2・d−1) where coral bleaching occurred and zooxanthellae counts in Acropora formosa reached their lowest concentrations on 13th February (Figure 4(f)). Whilst high levels of LLC in the summer can lower solar radiation levels and SSTs thus protecting coral reefs from the full effects of high solar radiation levels [23] [24] optically thin LLC can also increase solar radiation levels in the GBR [47] [48] .
For example, elevated LLC (>4 oktas) can clearly attenuate solar radiation (Figure 10), but extreme solar radiation levels (~30 MJ・m−2・d−1) occurred in the local region at both low (1 - 2 oktas) and medium (3 - 4 oktas) levels of LLC (see circled area in Figure 10), suggesting a positive radiative effect from low and medium levels of LLC as reported for this region [47] . From November to early January over a period of 29 days solar radiation levels were often extreme (>29 MJ・m−2・d−1). From 19th-28th December solar radiation levels often ranged from 29 - 30 MJ・m−2・d−1, contributing to the second highest DHW of +2.31˚C in Magnetic Island reef flat waters (Figure 7), with SSTs ranging from 27.33˚C - 31.76˚C, strongly suggesting that coral bleaching could have occurred earlier than the first week of January. This suggestion is supported by the extremely low levels of zooxanthellae in Acropora formosa at Nelly Bay from 6th November to 16th December (1.2 - 1.4 × 105 zooxanthellae polyp−1), compared with zooxanthellae counts in recovering Pocillopera damicornis colonies of 2.67 × 106 at the same location in July [31] . Bleaching of A. formosa in early January further reduced zooxanthellae concentrations to 0.5 × 105 zooxanthellae polyp−1 on 13th February (Figure 4(f)). The solar radiation levels over Nelly Bay reef in January were exceptional and bleached the upper surfaces of Acropora coral at Nelly Bay reef [46] . These high levels of solar radiation undoubtedly contributed to the very high SSTs of 30.5˚C - 33.3˚C at Nelly Bay in the first week of January [41] [44] and most probably led to the corals bleaching more severely, with corals continuing to loose zooxanthellae right up to mid-February (Figure 4(f)). However, directly after the bleaching from the 10th-15th January Magnetic Island surface waters cooled significantly with a DCW of −2.66˚C (Figure 7), most probably aided by the high levels of LLC (Figure 6(c)), which periodically reached 4 - 6 oktas (mean of 3 hourly observations) and produced 64 mm of rainfall (Figure 3(c)). From 19th-23rd January, daily air temperatures included a series of unusually high minimum daily temperatures, including the second highest minimum air temperature (29˚C) recorded locally since 1942 at the time and which coincided with increasing levels of LLC during the nighttime (Figure 6(c)) [44] . Daily air temperatures then fell sharply from 29.3˚C on 24th January, to 24.8˚C on 31st January 1994, aided by very high levels of LLC from 21st January to 3rd February (Figure 6(c), Figure 6(d)).
As solar radiation levels increased so did DMSw and DMS flux and were significantly and directly correlated with each other (see Table 1), possibly reflecting increased photosynthesis of reef organisms that produce DMS [28] [49] , and/or effects of SST and tidal stress that releases DMSw and DMSa from coral reefs [10] [20] [33] . Production of coral mucus enriches reefs with extremely high DMS concentrations, particularly when stressed [34] [49] , and together with other volatile organic substances such as isoprene from corals [50] can rapidly enhance the oxidation of DMSa. As solar radiation levels increased over coral reefs so did DMSw and DMS flux and explains why there is such a good correlation of DMSw with increasing SST for Pioneer Bay reef (see Table 1). However, no such correlation occurred for Nelly Bay reef when SSTs > 30˚C were included. When SSTs > 30˚C were excluded a significant DMS-SST correlation was also observed for Nelly Bay reef (Table 1). ![]() concentrations were also significantly positively correlated with increasing SSTs and significantly negatively correlated with DMSw (see Table 1) as previously reported [1] . The huge increase of
concentrations were also significantly positively correlated with increasing SSTs and significantly negatively correlated with DMSw (see Table 1) as previously reported [1] . The huge increase of ![]() concentrations from January to February often coincided with high sporadic fluxes of DMS from Nelly Bay and Pioneer Bay reefs (Figure 4(b), Figure 4(e)) and could explain the high (NH4)2SO4 concentrations (sulfate aerosol) that occur periodically in marine air masses over healthy coral reefs in the GBR [26] . This evidence strongly suggests there may be a link between DMSa emitted from coral reefs in the GBR, sulfate aerosol particles from oxidation of DMSa [26] , and the elevated levels of condensation nuclei (CN) measured over the GBR [51] which are precursors of CCN and LLC (Figure 1).
concentrations from January to February often coincided with high sporadic fluxes of DMS from Nelly Bay and Pioneer Bay reefs (Figure 4(b), Figure 4(e)) and could explain the high (NH4)2SO4 concentrations (sulfate aerosol) that occur periodically in marine air masses over healthy coral reefs in the GBR [26] . This evidence strongly suggests there may be a link between DMSa emitted from coral reefs in the GBR, sulfate aerosol particles from oxidation of DMSa [26] , and the elevated levels of condensation nuclei (CN) measured over the GBR [51] which are precursors of CCN and LLC (Figure 1).
The chemical composition of CCN is predominantly sulfate neutralized to varying degrees by ammonium [52] . Once a cloud droplet is formed it will scavenge ammonia (Liss and Galloway, 1993 [53] ). This production of ![]() is vital to the formation of sulfate aerosol since it charge balances the sulfate in the atmosphere (Figure 1). Much of the light scattering properties of sulfate aerosol has a range of compositions from H2SO4 to (NH4)2SO4, which can enhance solar radiation levels [7] . The interaction of the sulfur and reduced nitrogen cycles in the atmosphere over the GBR can influence the productivity of the GBR. The more sulfate aerosols present in the atmosphere, the greater the rate of nitrogen deposition due to scavenging of ammonia by sulfuric acid. In this way sulfate aerosols can essentially act as a carrier of nutrient nitrogen into GBR waters which could be greater than land-inputs of nitrogen [53] . Further studies are urgently needed to quantify this atmospheric nitrogen and sulfur input to the GBR [J. Ivey; PhD studies].
is vital to the formation of sulfate aerosol since it charge balances the sulfate in the atmosphere (Figure 1). Much of the light scattering properties of sulfate aerosol has a range of compositions from H2SO4 to (NH4)2SO4, which can enhance solar radiation levels [7] . The interaction of the sulfur and reduced nitrogen cycles in the atmosphere over the GBR can influence the productivity of the GBR. The more sulfate aerosols present in the atmosphere, the greater the rate of nitrogen deposition due to scavenging of ammonia by sulfuric acid. In this way sulfate aerosols can essentially act as a carrier of nutrient nitrogen into GBR waters which could be greater than land-inputs of nitrogen [53] . Further studies are urgently needed to quantify this atmospheric nitrogen and sulfur input to the GBR [J. Ivey; PhD studies].
Continuous SSTs measurements for Magnetic Island reef flat waters revealed various heating and cooling periods, interspersed with stable SSTs (Figure 7). The most extreme heating occurred in late December and early February when solar radiation levels were extreme and LLC was high (Figures 3(a)-(c)). Most cooling periods (negative climate feedback) occurred during low tides (<0.8 m) and decreasing DMS fluxes (Figure 8(b), Figure 8(c)), whilst most heating periods were often associated with higher low tides (>0.8 m) and elevated DMS fluxes, solar radiation and LLC (Figure 8(b), Figure 8(c)). Alternation between these two states or types of feedback (cooling and warming) is indicated in this research (Figure 8(a), Figure 8(b), Figure 9). A cooling period (period 4 in Figure 5) of DCW −1.4˚C, associated with a low tide of 0.8 m coincided with high levels of LLC and rainfall of 163 mm during the 23rd January to 3rd February period (Figure 3(d)). The combination of low spring tides and rainfall on the aerially exposed reef flat could result in extremely high DMS fluxes from the two coral reefs and could explain the high levels of LLC over reefs in the region from 23rd January to 3rd February (Figures 6(c)-(d)), causing lowered SSTs. In our study the cooling and heating periods identified were significantly correlated with DMS fluxes from the two reefs (p < 0.10) and suggest that as the flux of DMS from coral reefs increases cooling of reef flat waters occurs (Figure 7, Figure 8(c)). This could reflect increases in reef-generated DMSa and oxidation to sulfate aerosol and CCN formation (Modini et al. 2009), leading to enhanced LLC and periodic rainfall (Figure 1). Whilst detailed atmospheric measurements still need to be carried out over the GBR to more fully understand this climate feedback there is now more evidence that SST cooling by LLC does in fact occur in the wider GBR [18] [20] [23] , thus implicating reef emissions of DMSa.
This cooling trend in the central GBR has been examined across all study summers and shelf positions [23] . Leahy et al. found SSTs exhibited distinct drops during periods of high cloud cover, and conversely SST increases during periods of low cloud cover, with a 3-day temporal lag between a change in cloud cover and a subsequent change in SST. The relationship was strongest in both El Nino (2005) and La Nina (2008) study summers, at the inner-shelf position in those study summers, where the LLC cover tends to accumulate [23] . Interestingly these authors found occasions when SSTs decreased by ~0.5˚C - 1.5˚C during cloud cover increases (January and March), and situations where SSTs increased (February), a situation we have found in our study.
In our study different LLC-SST correlations occurred for different cooling and heating periods. For cooling periods in late November, mid-December, and almost all of March (periods not affected by coral bleaching), increasing levels of LLC were significantly correlated (p < 0.001) with decreases in SSTs (Figure 9(a)). However, in mid-January very high SSTs ranging from 31˚C - 33˚C occurred in Magnetic Island reef flat seawater and coincided with very low levels of LLC cover (<2.5 oktas) (Figure 4(c)). For a cooling period from 23rd January to 3rd February, high levels of LLC (Figure 4(c), Figure 4(d)) reduced SSTs to below 30˚C (Figure 5), coinciding with enhanced DMS fluxes from Pioneer Bay reef within this period (Figure 4(b), Figure 4(d)). During almost all heating periods two correlations again occurred between LLC and SSTs. For example, for heating periods that did not include the coral bleaching event in the first week of January (i.e. periods 1, 2, 3, 5, 6, 7) (Figure 9(c)), increasing levels of LLC were accompanied by increases in SSTs. For heating periods 1, 2, 3, 4, 7 (which included the coral bleaching period 4 from 2-9th January) decreases in LLC were accompanied by increases in SSTs as one might expect (Figure 9(d)). It is thought that these contrasting effects on the LLC-SST correlation reflect the high solar radiation levels that can occur when LLC contains high concentrations of small diameter (nm) sulfate aerosol particles [26] , generated by the oxidation of DMSa produced from the reefs during low tides [28] [29] . These increases in DMSa have been observed as spikes or pulses, lasting from ~1 to 8.5 h, at Heron Island coral cay in the southern GBR [10] . These researchers have also observed that when convective rain falls onto exposed coral reefs in the GBR at low tides even higher spikes of DMSa can occur over reefs, possibly caused by intracellular changes in DMSP in corals [31] . It is thought that these natural stress events (low tides, high SSTs, rainfall) cause the coral to utilize intracellular DMSP to cope with the resultant osmotic, solar radiation and temperature stresses [10] [31] [32] [33] , and in the process produce enhanced levels of seawater and atmospheric DMS. Swan et al., [10] [50] have proposed that CCN produced from coral reef-derived DMSa oxidation products could contribute to cloud formation in the southern GBR as part of the convective internal boundary layer (CIBL) produced over Heron Reef [54] . These sporadic spikes of enhanced fluxes of DMSa occurred intermittently over the Heron Reef and were 2 to 30 times the DMS flux from the adjacent ocean surrounding the coral reef [10] [55] . According to Swan et al., [10] the island cay could assist the convective upward movement of DMS and its oxidation products via the CIBL to form localized convection clouds which may precipitate over the island and surrounding reef. Pro- cesses such as this could generate the proposed coral reef climate feedback described in this paper. Such a process would lower reef flat SSTs, which has been observed in this study at Magnetic Island.
In contrast, the high SSTs that accompany low or medium levels of LLC, could reflect increased scattering of solar radiation from a build-up of these small diameter sulfate aerosols in the marine boundary layer [7] [18] and could explain the presence of optically thin clouds that are often observed during coral bleaching in the GBR (see Figure 6 in Elvidge et al. [56] ). Changes in nss-sulfate aerosol number and LLC cloud amount could increase the long-wave and short-wave cloud radiative forcing [57] over coral reefs in the GBR, as suggested by the good correlations between DMSw and DMS flux and solar radiation (see Table 1), and could exacerbate coral bleaching events from this increased scattering of solar radiation. The emission of aerosol particles from the 1,000 km length of reefs in the GBR has been calculated to be 1019 particles s−1, comparable with emissions from land surfaces and forest fires [51] . These aerosol particles over the GBR consist of 60% ammonium sulfate and 40% non-DMS volatile organics, oxidation products of DMSa [26] . The results of Modini et al. [26] are the first direct observation of aerosol particle formation over the GBR with exceptionally high growth rates and very high aerosol particle concentrations. In light of our recent research these high aerosol concentrations could clearly reflect enhanced levels of DMSa emitted from coral reefs in the GBR during low tides [10] [28] [29] . Oxidation of DMSa to sulfate aerosols then occurs (Figure 1) and these aerosols grow to form CN [51] , precursors of CCN, potentially leading to increases in LLC cover which could increase net downwelling solar radiation and SSTs in the GBR [23] [54] [57] , causing cooling and warming of SSTs during the summer season.
An important parameter that governs the relative scattering and absorption of solar radiation and thus the net downwelling solar radiation is the single scattering albedo (SSA), which refers to the ratio of scattering to the sum of scattering and absorption [58] [59] . Aerosols have a net negative surface-cooling effect when SSA < 0.95, and a net positive surface warming effect when SSA > 0.95 [59] [60] . The net effect can change from a negative to larger positive forcing depending on the cloud fraction, surface albedo, and vertical aerosol concentration and cloud distribution [59] . Aerosols, in particular sulfates (from the oxidation of DMS, Figure 1), larger than ~0.05 µm provide the nuclei for most cloud droplets. Cloud droplets are the most important factor controlling the albedo of clouds and hence the temperature of our planet. Newly formed CN scatter radiation, but when they grow larger, acting as CCN (5 - 10 µm), their scattering cross-section increases 107. Under these conditions channeling effects occur where direct radiation is reflected by the clouds and solar UV radiation is enhanced. Such a situation has been observed in the Townsville region of the central GBR [48] , in the wider GBR [61] and during coral bleaching at Keppel Islands and Heron Island in the central and southern GBR [54] [56] , and could have accentuated the mass coral bleaching events in the GBR in 1998, 2002, and 2016 [40] . Low tides have a major and variable impact on emissions of DMSa from coral reefs in the GBR [10] [28] [55] with oxidation forming variable sulfate aerosol concentrations that would affect the net downwelling solar radiation [44] [54] . Under light winds and low tides in summer at the Heron Island reef flat, daily net downwelling shortwave solar radiation can exceed +800 W・m−2; with up to 95% of the net radiation during the morning heating the water column and benthic coral cover [54] . These extreme solar radiation levels caused heating of the reef which was exacerbated by a mid-afternoon low tide when the shallow reef water reached 34˚C and near-bottom temperatures 33˚C, exceeding the thermal tolerance of corals at Heron Island, causing coral bleaching [54] . Enhanced scattering of solar radiation from nss-sulfate aerosols derived from oxidation of DMSa produced from coral reefs, and associated cloud radiative effects, may have a bearing on the severity of coral bleaching events in the GBR [20] [63] and may reflect variation in the liquid water depth of the aerosols [62] . This net radiative forcing from clouds can be as high as four times as large as the radiative forcing from a doubling of CO2 levels in the atmosphere [57] and needs to be taken into account when ascribing coral bleaching events in the GBR solely to GHG warming.
6. Conclusion
Our research has provided preliminary evidence suggesting that variations in the flux of DMSa from coral reefs in the GBR may enhance the scattering of solar radiation from nss-sulfate aerosols derived from oxidation of DMSa, which could be responsible for the extreme solar radiation and SSTs observed during mass coral bleaching events. Changes in the LLC-SST relationship for the Magnetic Island fringing coral reef could indicate that the cooling and warming effects observed at this reef in summer may be due to changes in cloud radiative effects in the solar UV and thermal IR affecting downwelling radiation to the coral reef. These cloud radiative effects, as suggested by the different LLC-SST correlations, may have a bearing on the increased frequency of mass coral bleaching events we are now seeing in the GBR as SSTs increase from global warming [37] [40] [63] . Global warming in combination with cloud radiative effects from aerosols produced from the coral reef marine environment may partly explain the mass coral bleaching in February 2016 when one third of coral reefs in the northern GBR experienced severe bleaching decimating hundreds of coral reefs. Clearly more in-depth studies are needed in the GBR on this coral reef-climate feedback in order to critically assess the factors that affect cooling and warming of inshore GBR waters from aerosols produced from the coral reefs and how this may affect the climate of northern Australia and the western Pacific.
Acknowledgements
Thanks are extended to the staff of the Orpheus Island research station for their assistance and to Advan Muslim who collected samples from Nelly Bay reef. Dr Ray Berkelmans (AIMS) is thanked for providing the SST data from Magnetic Island. The completion of this research was assisted by a grant to Associate Professor Graham Jones by ARC Discovery Project grant (ARCDP) DP150101649 with Professor Zoran Ristovski of the Queensland Institute of Technology.