Received 31 December 2015; accepted 23 January 2016; published 26 January 2016

1. Introduction
Aspergillus Section Flavi is the major group of fungi associated with aflatoxin contamination in several agricultural commodities. Three species of this section are known to produce aflatoxins: A. flavus, A. parasiticus and A. nomius, with the first two important in the colonization of maize, which is a staple food in Kenya. Sclerotia are important perennating structures of fungi, especially pathogenic fungi as they remain dormant during stressful environmental conditions like drought and germinate when more favourable conditions, usually adequate moisture and temperature, arise. Fungi also sequester a number of secondary metabolites in sclerotia that act as a chemical defence system against predators [1] . Ability to form sclerotia and formation of metabolites has been associated with virulence in fungi [2] . The small sclerotia forming strain of A. flavus is the cause of the reported aflatoxicosis outbreaks in Kenya and has been isolated mainly in the Eastern Province of Kenya. The large sclerotia formers are predominant in the western part of the country and have been demonstrated to be able of producing large amounts of toxins in vivo and to cause chronic exposure to humans in this region [3] . The small sclerotia Aspergillus produce relatively more sclerotia and toxins compared with the large sclerotia formers. Erlich and Mark [4] reported that the timing of expression for some of the gene clusters for secondary metabolism in Aspergillus flavus was coordinated with sclerotial production and that the associated metabolites accumulated preferentially in sclerotia. This study explored the possibility of using the ability to form sclerotia as indication of toxigenicity among the large sclerotia formers isolated from maize in Nandi County, Kenya. Aspergillus flavus isolates were analyzed for the presence of two aflatoxin biosynthesis genes in relation to their capability to produce aflatoxins, targeting the structural genes aflD, and aflQ genes. The aflD gene encodes an enzyme that catalyzes the conversion of the first stable aflatoxin biosynthesis intermediate, norsolorinic acid to averantin [5] while the aflQ gene is involved in the conversion of O-methylsterigmatocystin (omst) to aflatoxin B1 (AFB1) and aflatoxin G1 (AFG1) and dihydro-O-methylsterigmatocystin (dmdhst) to aflatoxin B2 (AFB2) and aflatoxin G2 (AFG2) [6] .
2. Materials and Methods
2.1. Description of the Study Site and Sampling
The study was conducted in Nandi County, Rift Valley Province, which is the major maize growing area in Kenya. Nandi lies at latitudes 0˚ and 0˚34" North and longitude 34˚44" and 35˚25" East and covers an area of 2873 km2. The highest levels are 1800 m above sea level while the lowest stand at 950 m above sea level. It has bimodal rainfall distribution pattern, long rains received between March to June while short rains in September to November and one maize planting season of March to April. The average temperature is 20˚C with the highest recorded in the months of December to January (23˚C) and the lowest of 12˚C in the months of July to August [7] .
The study was part of a larger project whose aim was to study aflatoxins in the milk value chain. The selection of the locations for study in the project was based on rearing of dairy cattle and maize cultivation. The locations were Kaptumo in Nandi South, Kilibwoni in Nandi Central and Laboret in Nandi North. The required sample size was calculated using the formula described by [8] resulting into 225 households. Quota sampling was used to obtain the required sample size proportionately from each of the three locations. Households that participated in the study were selected randomly from the sample size of each location. The survey was conducted from August to December 2010.
Five hundred grams of shelled maize kernels were collected randomly in sterile brown paper bags from the households’ storage facilities, sealed and stored at 4˚C. In household stores with less than ten sacks, all sacks were sampled while for stores with more than eleven sacks, ten were randomly sampled [9] . Maize kernels were sampled from different parts of the sacks using a closed spear driven through the top and sides of each sack to obtain a total of 500 g of incremental samples.
2.2. Isolation and Identification of Aspergillus sp. from Maize Kernels
Five kernels were surface sterilized for 1 min in 2.5% NaOCl, washed in three changes of sterile distilled water and plated on ¼ PDA prepared from Potato Detrose Broth (Difco) and amended with 2 ml/L of lactic acid to inhibit bacterial contamination. Six replicates from each household’s samples were plated and incubated at 31˚C for 3 days. Fungal growth colonies on maize kernels were visualized using a stereo-binocular microscope (Magnus M24). Aspergillus colonies were counted and identified according to Pitt and Hocking [10] . Pure colonies of Aspergillus were obtained after single sporing and transferred onto PDA medium to study the macro and micro morphological characteristics for identification using the taxonomic keys of Klich [11] . The cultures from PDA were sub cultured onto Aspergillus flavus parasiticus agar (AFPA) as described by Pitt, et al., [12] and incubated in the dark for 42 - 72 hours at 28˚C to confirm group identification by colony reverse colour.
2.3. Sclerotia Formation by Aspergillus Isolates
A. flavus isolates recovered from maize kernels were tested for their ability to form sclerotia on different growth media. An aliquot of spore suspension containing approximately 106 conidia/ml was spread onto plates containing CzapekDox Agar (CZA), Czapek Yeast Extract Agar (CYA), Potato Dextrose Agar (PDA) and V-8 juices agar (V8) medium. This was done in four replicates and incubated for two weeks at 30˚C in darkness [13] . A sample of 100 sclerotia from each media (25 per Petri dish) was measured using a reticle in a 10X Nikon Stereo Miscroscope (Nikon Inc. Garden City. NY, USA) connected to a monitor. Eight plugs of 1 × 1 cm were taken from each plate and transferred to a clean plate containing 5 ml of 100% ethanol and sclerotia scraped from the plugs in a level 2 safety cabinet to reduce risk of human exposure. The sclerotia were counted.
2.4. Molecular Characterization of Toxigenic Potential of Aspergillus flavus Isolates
A spatula full of spores was transferred from a 7-day-old PDA culture and re-suspended in nuclease free water and total DNA extracted using a ZR Fungal/Bacteria DNA MiniPrep Kit (Zymo research, South Africa) according to the manufacturer’s instructions. The PCR amplifications were performed using the Dream Taq Master mix (Thermo Scientific). The amplification reactions were carried out in volumes of 50 µl containing; 25 μL DreamTaq Green PCR Master mix, 1 μM of each primer i.e. the forward primer and the reverse primer, 1μg of the template DNA and 23 μL of nuclease free water. PCR was carried out as follows: (1) 1 step at 94˚C for 3 minutes (2) 30 cycles of the following 3 steps 1 minute at 57˚C, (2) 1 minute at 72˚C and 3) one final 10 minutes step at 72˚C. Amplification products were visualized in 1.2% agarose gels stained with ethidium bromide. The gel was photographed under UV light at 254 nm. The aflD gene was tested using the primers Nor 1-F (5’-ACC GCT ACG CCG GCA CTC TCG GCA C-3’) and Nor 1-R (5’-GTT GGC CGC CAG CTT CGA CAC TCC G-3’) developed by Rodrigues et al., [14] . The aflQ gene was tested using the primers Ord 1-gF (5’ TTA AGG CAG CGG AAT ACA AG-3’) and ord-gR (5’GAC GCC CAA AGC CGA ACA CAA A-3’) [15] .
2.5. Aflatoxins Analysis in Vivo
Fifty isolates of A. flavus that were positive for either aflD or aflQ or both were grown on Yeast Extract Sucrose (YES) Agar [14] to test their aflatoxin production profile. The isolates were inoculated on 9 cm diameter Petri plates and incubated at 27˚C for 7 days in the dark. Aflatoxins were extracted using the methods described by Vega, et al., [16] and Smedsgaard [17] with some modifications. Using a 9 mm diameter sterile cork borer, 9 plugs were harvested uniformly from the plates into 50 ml propylene tubes. Three plugs were randomly picked and added to a 5ml amber screw-cap vial of known weight and weighed again. The plugs were extracted in 2 ml methanol: formic acid (25:1) for 1hour at 25˚C ultrasonically. Using sterile syringe, the extract was drawn from the vials and filtered through 0.2 µm nylon membrane filter discs into a clean auto sampler vial (11.6 mm OD × 32 mm height) and analyzed by LC/MS/MS (XevoTM Waters). Aflatoxin standards were supplied by Sigma (South Africa Part number 46300-U). A mix of aflatoxins contained 1000 µg/kg of AFG1 and AFB1 and 3000 µg/kg of AFB2 and AFG2. This mixture was then diluted 10×, 100× and 1000× to create 4 calibration standards (including the undiluted mix) (1 µg/kg, 10 µg/kg, 100 µg/kg, 1000 µg/kg). These 4 standards were injected separately onto the LC/MS/MS and a calibration curve was created for each of the 4 aflatoxins (peak area vs concentration). The injection volume was 10 µl for the aflatoxins standard mix and 5 µl for all the other standards and the samples. The sample concentrations were determined from the area of each of the aflatoxins using the calibration curve of each aflatoxin. The aflatoxins concentration in the Petri dishes was obtained using the equation described by Okoth et al. [3] .
2.6. Aflatoxins Analysis of Maize Samples
The maize samples on which fungal isolation was carried out as described above were screened for aflatoxin content using LC/MS/MS. Five grams of milled samples were each weighed into 50 ml sterile centrifuge tube. 20 ml extraction buffer was added to each sample. The tubes were shaken vigorously and placed in the incubator/shaker at an angle for 30 minutes at 25˚C and 200 rpm. The tubes were then transferred to Eppendorf centrifuge and the content centrifuged at 4˚C for 10 min at 500 rcf. A sterile syringe was used to remove ± 2 ml clear extract, which was then filtered through a 0.20 µm/nylon filterinto a 2 ml Eppendorf tube, kept in fridge overnight. The contents were centrifuged for 10 min at 14,000 rpm and 1,800 µl clear extract pipetted into 2 ml Eppendorf tube for LC/MS/MS analysis.
2.7. Data Analysis
Data on Aspergillus incidence, toxigenicity and aflatoxin presence were analyzed using SPSS version 16. Qualitative and quantitative binary Chi-square tests for equality of proportions (Pearsons’ Chi-square test with Yates’ continuity correction) were used to compare frequencies of occurrence of isolates and to show whether there was significant difference between toxigenic and non-toxigenic isolates of A. flavus [18] Differences in the presence of sclerotia among the growth media were evaluated for significance using Chi-square test. Differences in sclerotial diameter between two growth media, PDA and V-8 juice, were tested using Wilcoxon rank-sum test while the influence of media on time of onset of sclerotia formation was tested using Kruskal-Wallis test. Correlation was used to test the association between inherent ability of toxigenic isolates to produce aflatoxins in vivo and corresponding aflatoxins accumaletd on maize samples.
3. Results
3.1. Aspergillus Species on Household Maize in Nandi District
A total of 19 Aspergillus species were isolated from 81% of the maize kernels. Members of Section Flavi were the most dominant and A. flavus most common though coexistence was observed, Table 1.
3.2. Scelrotia Formation by Aspergillus flavus Isolates from Nandi
Sclerotia formed by Aspergillus isolates were L-type with diameter ranging from 900 to3100µm. All isolates formed sclerotia. However, growth media influenced sclerotia formation ( ), Table 2. The highest frequency was observed in V8 agar followed by PDA. All the isolates that formed sclerotia on any of the other four media also formed sclerotia on V8 agar. V8 agar produced more sclerotia followed by PDA. The number of sclerotia formed varied with the strain (
), Table 2. The highest frequency was observed in V8 agar followed by PDA. All the isolates that formed sclerotia on any of the other four media also formed sclerotia on V8 agar. V8 agar produced more sclerotia followed by PDA. The number of sclerotia formed varied with the strain (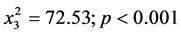 ) Time of onset of sclerotia formation varied significantly with strain and not media. While some isolates began forming sclerotia on the third day others
) Time of onset of sclerotia formation varied significantly with strain and not media. While some isolates began forming sclerotia on the third day others
![]()
Table 1. Aspergillus species isolated from household maize kernels in Nandi County.
![]()
Table 2. Influence of growth media on sclerotia formation.
*Average sclerotia score rating: 9 ≥ 901; 8 = 800 - 900; 7 = 700 - 799; 6 = 600 - 699; 5 = 500 - 599; 4 = 400 - 499; 3 = 300 - 399; 2 = 200 - 299; 1 = 1 - 199.
delayed until the seventh day though this variation was not significant (Kruskall-Wallis test; H = 2.58; df = 3; p > 0.05), Table 3.
Comparing the corresponding sclerotial sizes formed on V8 agar and PDA, revealed significant variations (Wilcoxon test, V = 224.5; p < 0.05). The same isolate produced larger sclerotia on V8 agar (1400 - 4000 µm) compared with PDA (950 - 2800 µm).
![]()
Table 3. Sclerotia formation on different media.
3.3. Toxigenic Potential of Aspergillus Isolates
Fifty nine percent of the isolates were positive for the toxigenic genes. Sixty seven percent of the isolates expressed only aflD gene while 31% expressed both aflQ & aflD, Welch two sample t-test revealed no evidence of variation in toxin levels in vitro between the isolates that expressed single bands (3082.77) and the isolates that expressed both bands (12723.35) (t10.624 = 1.3245, p = 0.2131), Table 4.
There was no correlation between the amount of aflatoxins produced in vitro when the isolates were grown on YESA compared with aflatoxins accumulated in corresponding maize samples (r = −0.006 and of p = 0.970), Figure 1.
![]()
![]()
Table 4. Aflatoxigenic gene profile of Aspergillus flavus and aflatoxin production in vitro.
![]()
Figure 1. Linear model showing relationship between aflatoxin produced in maize and those induced in vitro.
4. Discussion
A. flavus is the most common fungus occurring in maize in Nandi District. Co-occurrence of A. flavus with other species was observed and this has been demonstrated to reduce toxin production due to competition [19] . More than half of A. flavus isolates recovered from maize kernels in Nandi District formed sclerotia on media. V8 agar was the most favorable for sclerotia formation among the four media tested. V8 agar also showed less variation in sclerotia size and density as compared with the large variation observed among isolates grown on PDA. This confirms the ambiguity of the presence or abundance of sclerotia as a measure of strain toxicity. Isolates that did not produce sclerotia produced high levels of toxins and vice versa. Sclerotia are fungal perrenating structure but their presence or absence on culture media is not a reliable measure of potential of toxin production since media composition determines their formation. McAlpin [20] described that presence of different carbohydrates as well as to the amounts of these compounds in the agar medium induced formation of sclerotia initials and that interaction between the genome and physiological conditions affects sclerotia formation. The expression of gene depends on strain and the type and amount of nutrients in the medium. The disparity between aflatoxin production by the strains in vitro and in vivo confirms the strong dependency of biosynthesis of aflatoxins on fungal growth conditions.
Production of perennation structures in fungi and secondary metabolites are an adaptation to survival and so contribute to virulence index of the organism. Aflatoxins have been reported in all parts of the fungal structures, both vegetative and sexual. This implies that production of these structures in commodities would increase toxin levels. Knowledge about these factors and their synchrony in influencing the development of the structures and toxin production would also contribute to postharvest control strategy for aflatoxin accumulation in substrates. Perng-Kuang et al. [21] reported that the genes encoding for sclerotia formation are the same as those of aflatoxin formation. Most isolates expressed aflD or both aflD and aflQ though this could not be related to amount of aflatoxins produced. However, Galla [22] and Fakruddin [23] studied seven aflatoxin biosynthetic genes and reported that the variability existing in populations can be useful for understanding the toxicological risk as well as the selection of biocontrol agents.
Different households practice varied postharvest methods and though all the household samples were contaminated with toxigenic strains of Aspergillus flavus and with varied virulence, this was not the determining factor of the toxin level of accumulation in the household samples. With all A. flavus strains having intrinsic capacity to produce aflatoxins, environmental factors play a key role in production of toxins and so should be the major target in postharvest control strategy. This is an important observation for Kenya considering the report that 70% of the maize are harvested already contaminated with the fungus [3] , immediate measure of environmental factors should come into control at harvest. Moisture is the most important environmental factor that influences growth of moulds and toxin production followed by temperature [24] . Immediate drying following harvest and maintaining of the low water activity during storage and transportation would prevent phenotypic aflatoxin accumulation. This also strengthens the need of all players in the value chain doing their work and being sensitized of their significant role in the loop.
5. Conclusions and Recommendation
Though the S strain produces more sclerotia and toxins than the L strain, within the S strains and the L strains, variations occur with even high toxins produced by non-sclerotia formers.
Co-occurrence of fungal strains within a grain, observed disparity in ability of the strains to produce aflatoxins in vivo, the known radial growth mode of fungi and influence of environmental factors in eventual phenotypic aflatoxin accumulation in a commodity, are important factors that contribute to the heterogeneous distribution of aflatoxin in a consignment. As a consequence, it poses methodological difficulties in quantification of the toxins. These are some of the factors that should be taken into consideration in developing postharvest control strategies. The importance of co-occurrence and competition is seen in the use of atoxigenic strains of A. flavus to control the toxigenic strains. Depending on the particular combination of external growth parameters, the biosynthesis of aflatoxin can either be completely inhibited or the pathway fully activated. Knowledge about these relationships will enable an assessment of which parameter combinations can control aflatoxin biosynthesis and which are conducive to phenotypic aflatoxin contamination.
Acknowledgements
This work was a part of the Safe Food Safe Dairy Project funded by the Finnish Ministry of Foreign Affairs.
Conflict of Interest
There is no conflict of interest from other co-others in the publication of this manuscript in this journal. All the co-others have contributed in the preparation of the manuscript up to the submission stage.
NOTES
![]()
*Corresponding author.