Dynamics and Activity of Sulfate-Reducing Bacterial Populations in Paddy Soil under Subsurface Drainage: Case Study of Kamboinse in Burkina Faso ()
1. Introduction
Sulfur is among the most abundant elements on the Earth [1] . In rice soils, S cycle has two major aspects: plant nutrition and sulfides toxicity to the plant. Rice growth requires an adequate supply of S, depending on variety and yield, thus a rice crop removes between 8 and 17 Kg S ha−1 from the soil [2] . Microorganisms play an important part in sulphur transformations. Sulfate is taken up as a nutrient and reduced to sulfide, which is then incorporated into sulphur-containing amino acids and enzymes [1] . Sulfate-Reducing Bacteria (SRB) are ubiquitous and quantitatively important members in many ecosystems [1] [3] . They constitute a group of prokaryotes with diverse morphological and metabological characteristics, able to use several low-molecolar organic compounds, including mono- and dicarboxylic aliphatic acids, alcohols, polar aromatic compounds and hydrocarbons [4] - [6] . SRB are involved in numerous reactions under anaerobic environments [7] -[9] . SRB also constitute a diverse group of prokaryotes with the common ability to metabolize oxidized sulfur compounds. They are universally distributed where sulfate reduction is the dominant biomineralization pathway as fens [10] - [12] , estuary sediments [13] - [15] . Rice paddy fields also represent a freshwater environment in which sulfate reduction occurs and from which sulfate reducers have been isolated [6] [12] [16] . SRB gain energy for cell synthesis and growth by coupling the oxidation of these organic compounds or molecular hydrogen (H2) to reduction of sulfate (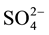 ) in sulfide (H2S, HS−) [1] [17] . Although most rice soils in the world are S-deficient, significant reduction of sulfate has been reported in a wide range of rice fields [2] [6] . Sulfate reduction is one of the most common and characteristic features shared by various types of paddy soils. It could be responsible to 95% - 97% of sulfides production in rice field [6] . Production of sulfides, at levels that cause rice injury, is observed in many countries, along with Burkina Faso where many rice fields soils are abandoned due to sulfides [18] [19] . Jacq [20] has described sulfides toxicity pathway in rice fields. By toxic sulfides production, SRB may be causative agents of limb discoloration. The high quantity of sulfides ions in the soil solution upsets the mineral element balance in the rice, affects its growth and causes crop yield losses depending on the sulfides concentration in the soil solution and the cultivar tolerance [21] [22] . Hydrogen sulfide may inhibit seed germination [20] , or causes early death of 30% - 100% of crop [23] . Up to now, sulfide toxicity in rice fields had been exclusively attributed to the activity of Sulfate-Reducing Bacteria [24] . The sulfide toxicity generally occurs in degraded paddy soils, poorly drained organic soils, and acid sulfate soils [21] .
) in sulfide (H2S, HS−) [1] [17] . Although most rice soils in the world are S-deficient, significant reduction of sulfate has been reported in a wide range of rice fields [2] [6] . Sulfate reduction is one of the most common and characteristic features shared by various types of paddy soils. It could be responsible to 95% - 97% of sulfides production in rice field [6] . Production of sulfides, at levels that cause rice injury, is observed in many countries, along with Burkina Faso where many rice fields soils are abandoned due to sulfides [18] [19] . Jacq [20] has described sulfides toxicity pathway in rice fields. By toxic sulfides production, SRB may be causative agents of limb discoloration. The high quantity of sulfides ions in the soil solution upsets the mineral element balance in the rice, affects its growth and causes crop yield losses depending on the sulfides concentration in the soil solution and the cultivar tolerance [21] [22] . Hydrogen sulfide may inhibit seed germination [20] , or causes early death of 30% - 100% of crop [23] . Up to now, sulfide toxicity in rice fields had been exclusively attributed to the activity of Sulfate-Reducing Bacteria [24] . The sulfide toxicity generally occurs in degraded paddy soils, poorly drained organic soils, and acid sulfate soils [21] .
For the experiment was developed to measure the effect of subsurface drainage on microbiological and chemical parameters sustaining sulfides toxicity in paddy fields and on rice yield. To eliminate excess sulfides in soil through water management, concrete microplots were constructed, filled with a clay-loam soil and water was drained periodically during rice, as described previously [25] . The Sulfate-Reducing Bacterial (SRB) populations’ density and sulfides content in the paddy soil were monitored during the cultural cycle of FKR 19 rice variety.
2. Materials and Methods
2.1. Expriment Site Location
The site for the experiments was located at Kamboinse in the central plateau of Burkina Faso (12˚26'48"N and 1˚33'45"W). Experiments were performed on the experimental site of the International Institute for Water and Environmental Engineering (2iE) from August to December 2011. The physical and chemical properties of the soil used in the experiments were described in our previous report [25] .
2.2. Climatic Characteristics of the Study Site
The site for experiments is located in the central plateau of Burkina Faso which supports five months of rainy season (Juin-October) with peaks from July to August, and seven months of dry season (November-Mai). During the period of study (August-December) thirty rainy days with an average rainfall of 284.8 mm were recorded (Figure 1). The average of daily temperature was 28˚C and ranged from 24.6˚C (minimum) to 30.9˚C (maximum) (Figure 2). The daily relative hygrometry (RH), varied from 39% to 78% (Figure 2), with a sunshine average of 7.26 hours/day (Figure 2).
2.3. Plant Material
For the experiment, FKR 19 rice variety (Farako-Bâ rice) was used. This variety originates from the Asian species Oryza sativa L (Japonica varietal group), and is adapted to the rainfed rice. The variety results from a cross- breeding performed by the Institute for Environment and Agricultural Research (INERA Farako-Bâ, Burkina Faso).With a cycle of seedling-maturity of 120 days, this rice variety has an average yield estimated to 5 - 6 tons/ha.
2.4. Microplots Experiments
Experiments were carried out as previously described by Otoidobiga et al. [25] and Keïta [26] . Twelve microplots
![]()
Figure 1. Monthly rainfall and monthly number of rainy days from August to December 2011.
![]()
Figure 2. Monthly average temperature and relative humidity from August to December 2011.
of 1.50 m × 1.20 m × 1.20 m in size (total internal volume: 1.31 m3) were constructed in reinforced concrete to prevent cracks due to temperature changes following the alternation of the rainy and the dry seasons. At the bottom of each microplot (1.45 m depth), perforated PVC pipes were installed and connected to an external tap through the concrete wall for sub-drain the soil (Figure 3). The design was intended to improve circulation of water and oxygen within the rice roots zone. Microplots were filled with the clay-loam soil described above [25] , and then flooded for 1 week. After the flooding period of soil, two 15-day-old plants of FKR 19 rice were transplanted at 20 cm hill intervals. Then, the plants were continuously flooded from rice transplanting until rice maturity and harvest. Three replications and four modes of drainage were performed throughout the study: without drainage (control: T) and drainage for 7 days (P1), 14 days (P2) and 21 days (P3), respectively by regulating the drained water flow from the bottom tap (Figure 3). Chemical fertilizers (NPK and urea) were applied in the microplots according to the recommended doses of 300 kg/ha for N-P-K (14 - 23 - 14) at transplanting and 100 - 150 kg/ha for urea in two further dressings (at rice transplanting, and 60 days after transplanting, respectively) [27] [28] .
2.5. Enumeration of Sulfate-Reducing Bacterial Populations
Populations of Sulfate-Reducing Bacteria were enumerated in microplots during the rice growth stages by the Most Probable Numbers methods using lactate and sulfate as substrates. The basal medium for enumeration was the medium described by Dianou and Traoré [19] .The medium composition was as follows (per liter of distilled water): [A] sodium lactate 60% (12 ml), K2HPO4 (0.35 g), KH2P04 (0.25 g), Na2S04 (4.00 g), MgS04, 7H2O (2.00 g), NaCl (0.5 g), NH4Cl (2.00 g), yeast extract (1.00 g), resazurin solution 0.1% (1 ml), 1 ml of trace elements solution [29] ; [B] Na2S, 9H2O (2.5 g), Chlorhydrate of cysteine (1.25 g), NaHCO3 (4.00 g). The heat-stable salts [A] solution was autoclaved and cooled under an atmosphere of N2. Then, the medium was supplemented with components [B] solution, to final concentration of 2%. The pH was adjusted to 7.2 by addition of NaOH 1N solution. Tubes were incubated at 37˚C for two weeks. Formation of colloidal CuS from total sulfides (H2S, S2− and HS−) according to Cor-Ruchwich [30] was used for detection of positive tubes after incubation period. The most probable numbers of SRB were calculated from a table of MPN for three tubes.
2.6. Determination of Sulfides Content in Soil
From the soil sampled for bacterial enumeration and at the same periods during the rice cultural cycle, the sulfides were extracted using extraction medium, according to the method adapted from Chaudhry and Cornfield [31] . The extraction medium composition was as follows (per litter of distilled water): CH3 (COO)2 Zn, 2H2O (50 g), CH3COONa, 3H20 (12.5 g). Cor-Ruschwich [30] method was then used to measure the content of sulfides in the soil solution. The extracted sulfides solution (2 ml) was removed by syringe and rapidly injected
![]()
Figure 3. Microplots design for the experiments [25] [26] .
into tubes containing 2 ml of a copper reagent. Immediately after mixing (for 5s), the absorbance is measured at 480 nm using a Spectronic 61 photometer.
2.7. Statistical Analysis
Data obtained were analyzed for SRB populations’ development and activity, drainage mode, rice growth stage and rice yield variations using the Student’s t-test and XLSTAT 7.5.2 statistical software. Mean parameters were compared according to the Newman Keuls’ test at 5% probability level.
3. Results and Discussion
3.1. Effect of Soil Moisture on SRB Populations Dynamic and Activity
In the present study, densities of SRB enumerated on soil before flooding, when the soil was dried and at transplanting day after soil flooding, are shown in Figure 4 and expressed as log 10 (most probable population number g−1 dry soil).
Sulfides content in soil before flooding when the soil was dried and at transplanting day after one week of soil flooding, are shown in Figure 5 and expressed as log 10 (µg/g dry soil). The concentration of sulfides increased after one week of flooding in the soil for all the microplots.
Rice paddy fields represent a freshwater environment from which sulfate reducers have been isolated and in which sulfate reduction occurs [1] [6] [16] [19] [31] . Many studies showed that sulfate-reducers are common in flooded soils and they are also found near rice roots [8] [32] .
Thus ours results are in agreement with those obtained by Dianou et al. [33] , who reported a population increase of 200%, 139%, 119%, 66%, 40%, 59%, 42% and 22% in the soils of Burkina Faso, after flooding. Garcia et al. [33] reported that SRB populations quantified in the paddy soils of Senegal were influenced by the redox potential related to flooding [34] . Indeed, soon after the flooding of the rice fields, O2 is removed from the bulk soil via respiratory processes, and large amounts of minerals and nutrients available for bacterial growth were released in the soil solution [16] [35] . In these anaerobiosis conditions the strictly anaerobic bacteria, including sulfate reducers, become active. Thus, significant reduction of sulfate occurs in a wide range of rice fields [16] .
![]()
Figure 4. Densities of Sulfate-Reducing Bacteria in soil before flooding, at transplanting day, and in soil near rice roots during the cultural cycle of FKR 19 rice in microplots without drainage (T), and drained for 7 days (P1), 14 days (P2) and 21 days (P3), respectively (means of 3 replicates).
![]()
Figure 5. Evolution of the soil sulfides content during the cultural cycle of FKR 19 rice in micro- plots not drained (T), and drained for 7 days (P1), 14 days (P2) and 21 days (P3), respectively (means of 3 replicates).
3.2. Effects of Rice Plant on SRB Populations’ Dynamic and Activity
The number of SRB in the soil near rice roots increased after one week of flooding for all microplots (Figure 4). An increase in SRB population number of 95.32%, 93.03%, 86.62% and 88.7% was recorded in the soil of the control (T), P1, P2 and P3 microplots, respectively. Our study also revealed that the number of SRB in soil near rice roots increased gradually with fluctuations from the transplanting day to rice flowering and maturity stages in all the paddy microplots (Figure 4). In most microplots, the highest densities of SRB were recorded from rice tillering and flowering to maturity stages, at which bacterial population number could reach 107 to 108/g dry soil (Figure 4).
The sulfides content in soil near rice roots also increased gradually with fluctuations during the rice cultural cycle and the highest levels were recorded from rice tillering and flowering to maturity stages at which it could reach 102 to 103 µg/g dry soil (Figure 5).
Thus, SRB population density and sulfides concentration in soil near rice roots appeared particularly high at the tillering flowering and maturity stages of rice growth (Figure 4 & Figure 5).
Wind et al. [36] who reported that the number of Sulfate-Reducing Bacteria were higher in planted than in unplanted rice microcosms support our findings. Many studies reported also that sulfate reduction rates and sulfate concentration were high at the root surface and near the rice roots [36] [37] .
Our results are also in agreement with those of Ouattara [18] and Dianou and Traoré [19] who reported that after rice transplanting the numbers of SRB increased significantly in the plant rhizosphere. Dianou and Traoré [19] reported also that populations increased gradually with fluctuations from transplanting day to rice flowering and maturity stages in the soil near rice roots during paddy plots experiments. Dianou and Traoré [19] and Jacq et al. [38] showed that bacterial population number could reach values of 107 to 109/g dry soil in most of paddy plot soils of Senegal and Burkina Faso, from rice tilling and flowering to maturity stages, respectively. The rice heading and repining periods which correspond to the highest level of reduced soil conditions in paddy flooded soil, may be conducive to the growth of SRB as underlined Dianou and Traoré [19] . Thus, the high physiological activity of rice plant at these stages may result in the production of more substrates available for bacterial growth [19] [20] [38] .
3.3. Impact of Drainage on SRB Populations’ Development and Activity
The variance of the SRB density in soil near rice roots, in relation to drainage and sampling period during rice growth is presented in Table 1. The Newman Keuls’ test revealed that the density of SRB in the paddy soil near rice roots was not significantly related to drainage (P = 0.187) (Table 1). Moreover, no significant difference was observed between the average numbers of SRB population in the soil of all the microplots (drained or not) (Table 2). However, the number of bacteria in the paddy soil near rice roots was significantly related to sampling period (P < 0.0001) and to combined both factors (P = 0.000, Table 1). These contrasted results could be partially explained by the MPN method used. Indeed, the analysis of bacterial communities by culturing methods, although valuable to characterize metabolic activities of their members, allows the characterization of only 1% to 3% of the microscopically-detectable cells in soils [39] [40] . Thus, molecular methods could be an indispensable tool to provide a more comprehensive description of the SRB community evolution in relation to drainage [3] [5] [40] [41] .
The variance of the soil sulfides content in relation to drainage and sampling period during rice growth is presented in Table 2. It appeared that sulfides content in the soil near rice roots was significantly related to drainage (P = 0.032) and to the sampling period (P < 0.0001). SRB have been traditionally considered as strict anaerobes [16] [42] . However, in the present study, the SRB population number in the soil of P1, P2 and P3 drained microplots increased slightly as compared to the control (18.45%, 1.06%, 8.76%, respectively) indicating that some among the SRB can survive and grow in aerobic compartments in the presence of low pressure of oxygen where a surplus of oxygen is released by drainage and by healthy roots [38] [43] .
These results are in agreement with the ones of Johnson et al. [44] and Dolla et al. [45] who found that some species of SRB were capable of slow linear aerobic growth in sulfate-containing medium in the presence of very low concentrations of oxygen. Thus, the abundance and metabolic activity of SRB in oxic zones of numerous biotopes are frequently evaluated as higher than those in neighboring anoxic zones [46] [47] . The high number of SRB found in these oxic environments indicates that these organisms are able to deal with temporarily exposures to elevated oxygen concentration [16] [48] . Wind et al. [36] show also that sulfate reduction also takes place on the roots of intact rice plants when O2 is allowed to diffuse to the roots through the aerenchyma system of the plants.
The density of SRB populations and the sulfides concentration in drained microplots could also be explained by heterogeneous distribution of oxygen through the drainage system leading to the formation of anoxic compartments [25] . A lack of oxygenation of such microsites in drained microplots may promote the SRB population survival [49] and the sulfides production [50] . Therefore, an efficient oxygenation of soil by water drainage would significantly reduce anaerobic SRB population’s development and activity.
![]()
Table 1. Variance of SRB number and sulfides content in soil near rice roots in relation to drainage and sampling period during the cultural cycle of FKR19 rice.
DF = degree of freedom; F = Fisher F; *significant P < 0.05; **significant P < 0.01; ns not significant P < 0.05.
![]()
Table 2. Effect of drainage on SRB population number in soil near rice roots during the cultural cycle of FKR19 rice in microplots not drained (T), and drained for 7 days (P1), 14 days (P2) and 21 days (P3) (means of 3 replicates).
Means with a same letter within a column are not significantly different according to Newman Keuls’ test P > 0.05.
3.4. Impact of Drainage on FKR 19 Rice Yield
In our study, symptoms of sulfides toxicity for rice based on IRRI standard evaluation system [51] weren’t evidenced during the microplot experiments. Although no significant difference (P = 0.209) was found for rice yield among treatments, drained microplots, in particular P2 ones showed the highest yield. Dobermann and Fairhurst [21] reported that an excessive concentration of hydrogen sulfide in the soil results in reduced nutrient uptake due to a decrease of root respiration. The typical symptoms linked to sulfide toxicity involves: rotting of roots, bronzing of leaves, poor growth at the reproductive phase, reduced ability to oxidize iron in the rhizosphere and poor yield [52] - [60] . The symptoms of sulfide toxicity can occur throughout the growth cycle of the rice [21] ; however no critical levels have been established to test sulfide toxicity. Our result can be explained by the chemical fertilizers (NPK and urea) applied in all the microplots. Indeed, according to Dobermann and Fairhurst [21] and Fageria et al. [61] , sulfides toxicity is generally associated with iron toxicity, which is defined as a multiple nutritive disorder further through excesses of H2S [61] . Deficiencies in minerals may enlarge the root permeability that enhances exudation, oxygen consumption and sulfate reduction, finally leading to severe sulfide toxicity [21] [61] - [63] . Many studies reported that sulfides toxicity for rice depends on nutrient supply; thus, amendment of paddy soil in mineral elements may reduce symptoms of sulfide toxicity for rice [64] [65] .
One raison of our results may be that the FKR 19 rice variety is not susceptible to sulfides toxicity as reported some studies [26] .
Rice plants can develop also physiological avoidance mechanisms to survive under toxic-sulfide condition [36] [49] . Indeed, sulfide toxicity depends on the strength of rice root oxidizing power, H2S concentration in the soil solution and root health [21] [38] .
Soil oxygenation by water drainage can affect sulfides profiles [49] [66] . The release of oxygen from the roots of rice causes radial redox gradients around the roots, leading to various chemical and microbial oxidation processes in the rhizosphere along with the oxidation of sulfide to sulfate [67] .
4. Conclusion
The study revealed important populations of cultivable Sulfate-Reducing Bacteria in soil near roots. The SRB populations and the production of sulfides appeared also impacted mainly by the soil flooding and by the physiological activity of rice plant at tillering, flowering and maturity stages. The effect of subsurface drainage on the above microbial dynamics and activity and the related consequence on rice production highlighted the highest rice yields for drained microplots. Although important concentrations of sulfides were recorded in the drained microplots, no deleterious effect was observed for rice development and rice yield. These findings suggested that the soil oxygenation through subsurface drainage might lead to chemical and microbial oxidation of toxic-sulfides to sulfate. Thus, an efficient oxygenation of soil by subsurface drainage may be a key factor of sulfides toxicity control in rice paddy field.
Acknowledgements
The authors would like to express profound gratitude to International Institute for Water and Environmental Engineering (2iE), CNRST/IRSS, General Direction Meteorology-Burkina Faso, International Foundation for Science, PACER-UEMOA/RABIOTECH, ISP-SUEDE/RABIOTECH, FCN-WAAPP and CNS-FL/WAAPP, for financial and technical supports.
NOTES
*Corresponding author.