Experimentally-Induced Inhibition of Growth in Melanoma Cell Cultures Separated by ~2 Kilometers When Both Share Excess Correlation Magnetic Fields: Macroscopic Evidence of Free-Space Quantum Teleportation? ()
1. Introduction
One of the most important possibilities for 21st century transposition of information within different localities is the technical production of reliable excess correlations without the direct involvement of an apparent medium. Whereas electromagnetic communication requires the assumptions of locality with spatial characteristics defined by magnetic permeability and electric permittivity from which the velocity of light can be derived, non-locality does not require an obvious medium. The occurrences of excess correlations of function between two non-tradi- tional localities, often described as “entanglement” [1] , are likely to involve multiple, different mechanisms. Traditional understanding of these processes have indicated this type of “entanglement” could only occur within subatomic and quantum domains. Here we report evidence that the survival of cancer cells in two distant loci display powerful correlations when the two spaces share specific types of circularly generated magnetic fields with changing angular velocities. The effect may be sufficient to develop practical applications for information “transfer” over distances through properties of photons for both electronic and living systems.
Quantum entanglement is a phenomenon whereby joint measurements of at least two systems exhibit stronger correlations than expected by classical explanations [2] . Quantum teleportation which involves the process of quantum communication has been demonstrated at distances that have ranged from 55 m to 16 km [3] . Hotta et al. [4] have calculated that quantum teleportation could occur without limitation by distance. Olson and Ralph [5] have shown that time-like entanglement could be extracted from a subcomponent of the quantum vacuum which has been can be interpreted in manner that is conducive to this process [6] [7] . As indicated by Vaziri et al. [8] the primary means for producing quantum communication for the foreseeable future will involve photons. According to most quantum approaches co-existence at some particular space-time coordinate is essential for the later display of entanglement properties, such as the reversal of polarity of one photon when the other’s polarity is altered. However Megidish [9] and his colleagues indicate that entanglement could occur between photons that never co-existed. Presumably this condition would require a shared third factor.
Julsgaard et al. [10] demonstrated experimentally that entanglement can occur between two macroscopic objects, specifically caesium gas samples containing about 1012 atoms. That macroscopic objects could display excess correlations at non-classical distances was further explored by Dotta et al. [11] . They based their design upon unpublished statistical analyses by Persinger and Lafreniere [12] who had noted that conjugate points in geomagnetic coordinates, when both exhibited properties of circular rotating fields within the larger angular momentum of the earth’s rotation, displayed evidence of “superimposition”. Dotta et al. [11] found that when two cultures of melanoma cells separated by 10 m were simultaneously exposed to circularly rotating magnetic fields the application of light flashes to one dish resulted in the emission photons as measured by photomultiplier tubes from the other. This “excess correlation” did not occur when the circularly rotating fields were not present.
Persinger and his colleagues [11] [13] developed an experimental protocol based upon the theoretical considerations by Tu et al. [14] that the photon displayed a non-zero rest mass. If this presumption were valid then the group and phase velocity of photons would be dissociated, thus creating additional vectorial features and properties of electromagnetic fields. We assumed that if the processes were isotropic their simulation could affect photon behaviour. The challenge was to build and to develop equipment that could create the conditions within a macroscopic context. Koren, Dotta and Persinger [15] devised a system by which electromagnetic fields with specific point durations generated by computer software could be generated around a circular array of eight solenoids. The rate of change of the circular angular velocity, which would technically be a second derivative, could be increased or decreased by the same software.
While discerning if photons themselves were the operational unity of the “excess correlations” between either cerebral or cellular activity [11] by measuring the non-local photon radiant flux density during chemoluminescent reactions (hydrogen peroxide and sodium hypochlorite solutions), Dotta and Persinger [13] identified one particular effective sequence. Doubling of the power densities of photon emissions occurred when both loci in which the chemoluminescent reactions occurred were first exposed to circularly moving magnetic field whose group velocity was accelerating and phase velocity decreasing and then to a circularly moving magnetic field whose group velocity was decelerating but phase velocity was increasing. In these instances the single injection of a reactant (hydrogen peroxide) into each locus of hypochlorite solutions resulted in a doubling of photon output as if the two loci were the same space and hence had been injected with twice the amount. The excess correlation was demonstrated at distances of 10 m and 3 km.
Clusters of cultured cells (~105 to 106) emit photons at radiant flux densities in the order of 10−12 W・m−2 [16] . The photons are more than by-products of metabolism and have been argued to be the primary mode of inter-cell communication [17] [18] . Melanoma cells emit copious amounts of photons during their habituation to room temperature after removal from incubation. The peak wavelengths of photons within the visible range shifted predictably as a function of the time since the removal from incubation [19] . There is direct and indirect evidence that these bursts of photons affect the activity of similar cells within their proximity. Drugs such as morphine elicit remarkably strong bursts of photons from malignant cell cultures [20] . In the present experiment novel equipment (toroids) was developed that captured the characteristics of the array of eight solenoids but was operated by easy-to-construct Arduino systems rather than the required custom constructed digital-to-analogue convertors (DACs) required in previous studies.
2. Materials and Methods
A total of 27 separate experiments (trials) were completed over a period of two months. B16-BL6 murine melanoma cells (~105) were cultured using 2.5 cc of DMEM culture media in 35 mm × 10 mm standard plastic dishes. For each experiment 2 plates of cells were placed in the center of the two toroids (1 plate per toroid), labeled as local or non-local. The exposure equipment and control systems are shown in Figure 1 and Figure 2. A schematic of the experimental design and circuit has been published elsewhere [21] . Each toroid was composed of a plastic ring whose diameter was 25.4 cm. It was wrapped with 225 turns of 16 gauge copper wire (for stereo speakers) and covered with black, vinyl electrical tape. The width between the bottom and top of the wire and plastic ring combined was 3.8 cm.
The local and non-local toroids were separated by either 3 m or 1.7 km (distance between the university lab
![]()
Figure 1. Pictures of the actual toroids and microcontroller system employed to generate the changing angular velocity pulsed magnetic fields.
![]()
Figure 2. Closer view of the micro-control (Arduino) system that operated the toroids.
and the local research hospital). The dish of cells was always placed in the middle of a solenoid at room temperature after being removed from standard incubation (37˚C). The toroid that was designated as local or non-local was counterbalanced within each treatment series. Local refers to the toroid within which the cells received small (50 μL) injections of hydrogen peroxide. Non-local referred to the toroid within which the cells never received any peroxide. Control plates (n = 2, per trial) were placed outside of the incubator for the same duration as those cells exposed to the experimental magnetic fields at distances where power meters indicated there was no discernable changes from the experimentally-generated magnetic fields.
Both pairs of plates containing cells were exposed to the excess correlation paradigm using decreasing and increasing phase (frequency) modulated magnetic fields. Their shapes have been published elsewhere [22] . The fields were controlled using an Arduino microcontroller system and laptop (Figure 2) similar to that described in more detail by Rouleau et al. [22] . By orienting the diode the current (fields) were rotated in the counterclockwise direction. Eight separate 3 ms pulses were generated in two patterns. For the decreasing frequency or phase modulating condition the 3 ms pulses (p) were generated in this manner: p, 20 ms, p, 22 ms, p, 24 ms, p, 26 ms, p, 28 ms, p, 30 ms, p, 32 ms, p, 34 ms. The sequence was then repeated for the duration that it was programmed. For the increasing frequency or phase modulating condition the 3 ms pulses were generated in this manner: p, 20 ms, p, 18 ms, p, 16 ms, p, 14 ms, p, 12 ms, p, 10 ms, p, 8 ms, p, 6 ms before the cycle repeated.
Consequently although the current within the copper wire was being generated continuously (and presumably moving ~108 m・s−1) there was a superimposed changing angular velocity implicitly associated with the different completion times for the generation for one sequence of the signals for decreasing (203 ms) and increasing (128 ms) angular velocity programs. The only additional component was an amplifier that was added to the circuit so that the strength of the field within the area occupied in the center of each toroid by the cells was 3 to 5 μT as measured directly by a power meter.
The paradigm was 12 minutes in duration. The first injection of 50 μL of either 300 μM, 800 μM, or 1 mM of hydrogen peroxide (diluted with culture media) was placed in the local dish 4 minutes after the initiation of the decreasing frequency modulated field. The pattern was then changed to an increasing phase (frequency) modulated field for 8 more minutes. Every 1 min for the next 5 minutes the same amount of hydrogen peroxide was injected into the local plate of cells. Hence there were a total of 6, 50 μL injections. After 12 minutes had elapsed form the activation of the fields, the Arduino systems for both loci were turned off and the cells were placed back into the incubators for 24 hours. After the 24 hours, cells from the plates were counted using the standard haemocytometer/Trypan protocol. The media containing the cells was centrifuged. The pellet containing the cells was then re-suspended in PBS (phosphate buffer solution). Within each chamber of the haemocytometer 15 μL of the solution was injected. The average (mean) of 8 grids was calculated and expressed in traditional formats (per 1.5 mL).
All experiments for local and non-local pairs for the distances of 3 m and 1.7 km were repeated five to six times. Only one experiment was completed per day. The experiments were also repeated when the excess correlation protocol was activated for both locations (separated by 1.7 km) for local and non-local reactions but only the volume (50 μL) of media (no hydrogen peroxide) was injected into the cells in order to control for injection artifacts. All results are presented as means and standard deviations (SDs) for each condition within the different experiments. Because of the conspicuous nature of the effects and the absence of overlap between the standard deviations we did not pursue traditional statistical analyses because all effects would be p < 0.01.
3. Results
The results were conspicuous and reliable. Figure 3 shows the means and standard deviations (SDs) for the cells remaining 24 hr after the serial (every min) injections of 50 μL of 800 μM hydrogen peroxide (local) into the dish and in the dish that did not receive any injections when both shared the same decreasing and increasing frequency (phase)-modulated magnetic fields delivered through different toroids separated by 3 m. Although one would expect the reduction of cell numbers in the cultures injected with the peroxide, the non-local cultures that were not injected but shared the changing angular velocities of magnetic fields displayed a reduction of cells that was approximately 50% (about 60 to 70 cells per 1.5 mL or about 1.2 × 105 cells) compared to the control cells (those that were removed from the incubator and placed near the local and non-local spaces but beyond the intensity of the fields. The difference between the local and non-local effect was about 10 cells per 1.5 mL or about 1 to 2 × 104 cells.
Figure 4 shows the means and standard deviations for the numbers of cells measured within the local and non-local plates 24 hr after the single exposure (12 min) to the excess correlation protocol when the toroids were separated by 1.7 km. The control conditions were dishes that were also removed from the incubators but not exposed to the excess correlation fields. Although the similarity of the cell effect for the cells exposed to the local condition would be expected because they were injected with the same amount of hydrogen peroxide, the cell loss within the non-local condition, even though the toroids were separated by 1.7 km rather than 3 m, was also
![]()
Figure 3. Means and standard deviations (for 5 experiments) for the Numbers of cells sampled from 15 μL of cell suspensions 24 hr after a single (12 min) exposure to the excess correlation rotational magnetic fields in pairs of toroids separated by 3 m. The local cells dishes were injected with hydrogen peroxide while the non-local was not. Control cells were not exposed to the paired fields or the hydrogen peroxide but had been removed from the incubator for the same duration.
![]()
Figure 4. Means and SDs (for 6 experiments) for the numbers of cells sampled from 15 μL 24 hr after a single (12 min) exposure to the excess correlation rotational magnetic fields in pairs of toroids separated by 1.7 km. The local cells dishes were injected with hydrogen peroxide while the non-local cells were not. Control cells were not exposed to the paired fields or the hydrogen peroxide but had been removed from the incubator of the same duration.
very similar.
Figure 5 shows the effects of injecting different concentrations of hydrogen peroxide (same unit volume of injection) into the local plates of cells. As expected increased concentrations of hydrogen peroxide was associated with more cell death in the injected (local) populations. However the non-local effect was non-linear. Compared to the dishes of cells from the control conditions, the non-local effect were marginally lower when the local cells were injected with either 300 μM or 1 mM of hydrogen peroxide. The maximum non-local effect in this experiment occurred when the concentration of peroxide injected into the local cells was 800 μM as was found in the previous experiments.
The means and SDs for the results of 5 experiments for numbers of cells within the control and local and non- local sites when only media was injected (no peroxide) are shown in Figure 6. There was no conspicuous suppression of cells numbers (cell loss) even when the toroidal fields were activated when just media was injected (rather than hydrogen peroxide) into the cells within the local condition. There was also no non-local effect.
4. Discussion
Direct and practical application of the variants of excess correlation will likely be one of the most important innovations for information superimposition between non-traditional distances in this century. Although the measurements of temporally congruent changes in properties of clusters of photons simultaneously in two different loci have been described as “macro-entanglement” or “teleportation” [4] , the essential operation is the superimposition of information in its most fundamental form. We are assuming, as has Vaziri et al. [8] that this fundamental form involves photons. Previous research has shown that excess correlation can occur over tens of meters to several kilometers for discrete photon emissions from controlled chemiluminescent reactions.
The results of the present experiments demonstrated that robust, reliable excess correlations occurred with a very specific sequence of changing angular velocities of discrete (3 ms) pulsed magnetic fields generated through double-circular geometry, i.e., a toroid. This unique configuration can be conceived as a three dimensional extension of a circle, a helical pattern, that is then contained within a circle. When the current generated with this double-circular geometry is pulsed, with the features of a “quantum well”, and the inter-pulse durations change systematically to simulate changing angular velocity (and momentum), the conditions are created to allow potentially Mach’s principle to be manifested as well as additional properties of photons if they display non- zero rest masses [14] .
The combination of Mach’s principle that emphasizes angular momentum [23] as an important contributor to
![]()
Figure 5. Means and SDs for the numbers of cells sampled from 15 μL 24 hr after they had been exposed to control conditions or after the local cells were injected with either 0.3, 0.8 or 1 mM of hydrogen peroxide. Note that when the local cells were completely eradicated by the hydrogen peroxide there was no evidence of excess correlation. The distance between local and non-local cell plates was 1.7 km.
![]()
Figure 6. Means and SDs for the numbers of melanoma cells after the serial 50 μL of media were injected into either the control or to the local and non- local dishes separated by 1.7 km.
the process by which a change in any subset of the universe affects the entire universe or set and the “extensional” properties of a photon may be one substrate for the occurrence of excess correlations. We have suggested that the specific duration of pulsed magnetic fields is derived from the largest and smallest features of the universe reflected by Hubble’s parameter and Planck’s constant. Persinger and Koren [24] had shown that the time required for the width of a proton to expand one Planck’s length would be about 3 ms. The required duration for an electron would be about 1 ms. Experimental evidence from photon emissions from chemo luminescent reactions supported this values [23] .
We designed the temporal parameters for this experiment to be very specific such that the 3 ms pulse would satisfy the proton condition. The differential between the addition or subtraction of 2 ms for the inter-pulse duration could be considered a persistent derivative (or rate of rate of change) that resulted in a beat frequency. This beat frequency or subtraction between the 3 ms and 2 ms durations would be 1 ms, or the predicted duration for the electron. If our model is correct the two basic components of matter, the proton and electron, and their spatial arrangements that determine molecular structure and activity and ultimately the properties of the living cell would be congruent.
Although the empirical support for such integration across quantum and macro-level processes will require more verification, the energy quantities that would be associated with the non-local effects reliably demonstrated in the present experiments can be tested by calculations. The median strength of the magnetic fields generated from the toroids within the center that was occupied by the plates of cells only once for 12 min was about 5 μT. According to the classic formula:
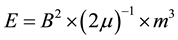 (1),
(1),
where B is the magnetic field strength, μ is the magnetic permeability (4π × 10−7 N・A−2) and m3 is the volume, the energy available within the volume of a melanoma cell with a diameter of 12 μm or a volume of 9 × 10−16 m3 would be ~10−20 J. This value is associated with the energy of forces over the distances between the potassium ions that are attributed to the resting membrane potential and the exchange of “information” between receptors and ligands [25] . This value is well within the quantity of energy associated with average force within the universe per Planck’s voxel (about 10−19 N) applied across the most ubiquitous wavelength of neutral hydrogen (21 cm) or 2.1 × 10−1 m [26] . Although interesting, the solution is not easily tested empirically.
However the potential bases for the narrow band of cell loss in non-local space that was associated with injection of the specific concentration of 800 μM of hydrogen peroxide into local space could be pursued quantitatively. The numbers of molecules injected during the protocol would have been 6 injections (5 × 10−5 L per injection) (8 × 10−4 M/L) (6.023 × 1023 molecules per M) or 1.45 × 1017 molecules. Our previous research has indicated that the proton, specifically the one associated with the hydronium ion, is a major mediator of the magnetic field effects. At pH 7.4, which would be the mode value of the cell culture as verified by the color of the media, the molarity would be 3.98 × 10−8. Hence the total number of protons involved with this total protocol would be 5.77 × 109.
This becomes important when the magnetic energy from the applied toroid fields is calculated for the total volume of hydrogen peroxide injected. According to Equation (1) for a 5 μT field the energy within 3 × 10−7 m3 (300 μL from 6, 50 μL injections) would be 3 × 10−12 J. This means that per second on average the synchronized energy from the two toroidal magnetic fields would be the quotient or 0.521 × 10−21 J. When applied over the total protocol duration of 12 min or 720 s, the total energy available to the system would be 3.67 × 10−19 J.
This quantity may be essential for understanding the process. When the value is divided by Planck’s constant (6.626 × 10−34 J・s) and this product is divided into the velocity of light in a vacuum, the wavelength is ~545 nm. This wavelength is within the visible range. In unpublished (and replicated) experiments we found that the average of the split spectrum of peak wavelengths of emissions of photons that were measured from melanoma cells during the 60 min period after they were exposed to other forms of pulsed magnetic fields was ~545 nm.
If a type of “experimental teleportation” [4] is involved with this protocol, then the properties of the electron should also be accommodated. The total numbers of cells in the re-suspended pellet in 1.5 mL of PBS, can be estimated quantitatively. Assuming an average of 130 cells in the control condition (e.g., Figure 3) which was from 8 fields the product multiplied by 18/8 (where 18 was the total number of fields occupied by 15 μL, the numbers of cells in 1.5 mL would be 2.3 × 105. The total number of cells multiplied by the unit energy per H+ from the peroxide reaction was 3.7 × 10−19 J for the total exposure. Hence the total energy for the system was 8.6 × 10−14 J. This is a particularly significant value because (when divided by the velocity of light squared) the mass equivalence is 9.5 × 10−31 kg, effectively the rest mass of an electron. For this value to occur very specific molarities of reactants and associated pH would be required as noted in our experiments. The convergence of the total energy within the system with a single electron classical mass could create the conditions for coherence [2] . It would also metaphorically become a type of “condensate” where by the state of the unit (electron) reflects the entire unit (the system).
Although the intricacies of such convergent reactions must still be isolated, these calculations may help explain the non-linear effect and why 800 μM of peroxide produced the greatest effects while 300 μM did not. For the latter the critical numbers of molecules were not available to produce the energy per molecule from the toroidal magnetic field to produce the energies that would be associated with the critical photon wavelength or the “condensate” of total energy of the entire population of cells. If all of the cells died from the peroxide, then there would be no source for the information to be mediated by the excess correlation.
The injection of the hydrogen peroxide into the cells is very likely to have promoted a variant of the well known reaction within hypochlorite ions that would have been available from the interaction of sodium and chloride with oxygen proximal to the cells’ plasma membranes. Our interpretation is that the emissions of these photons from the local cells were ultimately represented within the cells 3 m or 1.7 km away in the non-local spaces. Whether or not these photons occurred during the 12 minutes of exposure or later when cells from both the local and non-local sites were likely to have emitted photons because both had been exposed to the magnetic fields remains to be tested. The latter condition would suggest that once experimental “entanglement” had occurred the photon transpositions between spaces could occur during any subsequent time.
Another more information-based approach involves the quantum of energy associated with the dissipation of a bit of information into entropy. There could be the consequent transformation of virtual particles into matter determined by this information from the zero point vacuum oscillations into a non-local space [27] . Landauer’s solution derived from ln2 × kT where k is the Boltzmann constant and T is absolute temperature for the incubation temperature would be 4.2 × 10−21 J per bit. If we assume the total energy in the system from the magnetic field energy, as calculated previously was 8.6 × 10−14 J, then there would be ~2 × 107 bits available for the system. Because about 105 cells in the non-local space were eliminated, this would involve about 200 bits per cell. If this is sufficient to initiate a cascade that results in cell death must be verified experimentally.
These results are consistent with but not proof of our hypothesis that macro-excess correlation can be demonstrated in the laboratory. The narrow-band nature of the concentration of reactants that produced comparable effects at distances of 3 m and 1.7 km are also consistent with the superimposition of two loci into the same space such they behave as the same space. One interpretation of our results is that the excess correlation protocol produced the conditions by which the changes in protons and electrons within the local cells due to the peroxide injections resulted in transpositions of protons and electrons (or the virtual particles from which they originated) into both loci.
4. Conclusion
Specific sequences of changing angular velocities of serial point duration magnetic fields generated within toroids separated by 3 m or 1.7 km produced excess correlation from experimentally-induced changes in the viability of about 105 cells or the equivalent of 2 to 5 × 10−8 kg. One day after only 12 min of exposure to chemical reactions (local cells) that reduced cell numbers by 50% similar numbers of cells were reduced in plates of non- local cells but only if both local and non-local spaces shared the excess correlation protocol. The specific molarity of hydrogen peroxide that produced the maximum effect was associated with the numbers of protons in hydronium ions whose energy per molecule from the specific intensity of the magnetic fields would allow the conditions for photon emissions and production of the rest mass of an electron. Both phenomena may be manifestations of or means by which fundamental bits of information can be transposed in separate spaces. The results are consistent with transposition of the chemical reactions in the local cells to the non-local cells.
Acknowledgements
We thank Dr. W. E. Bosarge, Jr., CEO Capital Technologies for his support of novel technology and ideas. Thanks to Professor Nicolas Rouleau for lending his equipment and technical support and to Dr. Robert M. Lafrenie for the use of his laboratory space at the hospital.