Arsenic Induced Changes in Growth and Physiological Responses in Vigna radiata Seedling: Effect of Curcumin Interaction ()
1. Introduction
Arsenic (As) contamination in environment, from both anthropogenic and natural sources occurs is a global problem. As is a metalloid having properties of both metals and non-metals, and can undergo different ranges of chemical interactions in plants. Historically, the extensive use of arsenic-based pesticides has led to considerable contamination of domestic and agricultural land, through their use as lawn herbicides, and insecticides for rice plants, cotton, etc. [1] . Arsenic is a non-essential element and, in general, inorganic ones are phytotoxic. Arsenic is toxic to a wide range of organisms, including plants [2] . Studies on arsenic toxicity have shown that plant species not resistant to arsenic suffer considerable stress upon exposure, with symptoms ranging from inhibition of root growth to death [3] -[6] .
Exposure of plants to arsenic causes reduction in photosynthetic rate [7] , for example: in maize (Zea mays) plant, the rate of CO2 fixation in young plants treated with arsenic decreased by 20% and functional activity of PSII was reduced significantly [8] . The decreased photosynthetic rate under stress condition could be due to both stomatal and mesophyll limitations. The mesophyll factors could be of a different nature, such as disturbances in the chlorophyll, light and biochemical reactions from the Calvin Cycle [7] . In rice seedlings arsenic toxicity causes changes in carbohydrate metabolism leading to the accumulation of soluble sugars by altering enzyme activity. Sucrose synthase possibly plays a positive role in synthesis of sucrose under As-toxicity [9] .
Arsenic is known to induce oxidative stress in plants [10] , resulting in a range of responses in plants, including readjustment of transport and metabolic processes and growth inhibition in plants. As a result, the electron transport processes are impeded developing toxic systems. Several toxic reactive oxygen species (ROS) are generated in the cell wall region as well as inside the cell during the process, which affects membrane permeability, enzyme activity, metabolic pool, plant biomass, leaf chlorosis and necrosis [11] , probably due to the conversion of arsenate to arsenite, a process that usually occurs in plants by synthesizing both enzymatic and non-enzymatic antioxidants. Arsenate acts as a phosphate analogue and is transported across the plasma membrane via phosphate transport systems. Inside the cytoplasm it reacts with phosphate, replacing ATP to form ADP-As, precisely because of disruption of energy flow in cells [12] . On the other hand, Bertolero et al. [13] reported that arsenate reduced to arsenite in plant tissue, does not normally have enough cytoplasmic concentrations to exert toxicity. Arsenite reacts with―SH group of enzymes and proteins due to suppression of cellular function and death [14] . The conversion of arsenate to arsenite, in vitro observation, is brought about by glutathione [15] , with the concentration of high cellular plant tissues [16] .
Curcumin (1, 7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) is a polyphenol derived from the plant Curcuma longa. Extensive research over the last decades has indicated that curcumin exhibits anti-in- flammatory, anti-oxidant, anti-viral and anti-infectious activities [17] - [19] . Curcumin such as other polyphenols is as strong anti-oxidant [20] . It significantly decreases lipid peroxidation, regulates antioxidant enzymes and scavenges hyperglycemia-induced reactive oxygen species (ROS), [21] . Oxidative stress and inflammation are closely associated with tumor growth, [22] . All such effect of curcumin has been reported from animal system and no work has been done on the role of curcumin in plant growth. The present investigation is an attempt to understand the role of curcumin in heavy metal stress amelioration in plant in general and arsenic stress amelioration in mung bean in particular.
2. Materials and Methods
Mung seeds [Vigna radiata (L.) Wilezek] viability was carried out by the floatation method. The seeds obtained from the market were put in a beaker of water and allowed to stand for five to ten minutes. Seeds that sank were considered viable. Required amount of viable mung seeds were taken and surface sterilized with 0.1% HgCl2 solution for 3 - 5 minutes with successive shaking. After this, HgCl2 solution was thrown, washed thoroughly in tap water for 3 - 5 minutes, rinsed with distilled water for 2 - 3 times and decanted. Then the seeds were placed properly in petriplates containing moisten filter paper and germinated at 25˚C for 2 d. After two day of incubation the healthy germinated seeds with more or less equal height of shoots were transferred in the plastic cups (150 ml) containing half strength Hoagland nutrient medium and were grown under light in growth chamber for 5 days. After every two days the medium was changed for healthy growth. On the 5th day from the day of transfer, the plants were treated with 0, and 50 μM-cup was 50 μM, sodium arsenate solution. Each cup contains at least ten plants. In another set of treatments growing mung seedlings were subjected to different concentration of curcumin (0 μM, 25 μM, 50 μM & 100 μM) and their interaction with 50 μM sodium arsenate [(50 μM As + 25 μM curcumin), (50 μM As + 50 μM curcumin) and (50 μM As + 100 μM curcumin)]. The concentration of arsenic and curcumin used in this study was based on preliminary experiments. All these cups were kept under 16 h/8 h light/dark cycle in the growth chamber at 22˚C ± 3˚C. Plants were sampled after 48 h of teatments. The same treatments were used to test the effect of As, curcumin and their interaction at germination level. The viable mung seeds (25 in numbers) plated in petriplate containing water moistened filter is considered as control and in other set of treatments equal volume of treatment solution is used for germination of seeds. The appearance of the plumule at the filter paper surface was taken as germination. Germination percentage (%) was recorded after 48 h of incubation. After 48 h of treatment growing mung seedlings were sampled and root and shoot length were measured using centimeter ruler and were separated into root and shoot and then oven dried at 80˚C for 48 h to estimate the dry mass and expressed in g・plant−1. Root and shoot ratio was measured by dividing root length by shoot length.
Concentration of chlorophyll a (Chl a), chlorophyll b (Chl b), total chlorophyll and total carotenoid (Cx + c) concentrations were analyzed following Shabala et al., [23] and Lichtenthaler [24] methods, respectively and expressed as mg・g−1 FW. About 50 mg of leaf samples were placed in a glass vial, added with 5 ml of 95.5% acetone, and the glass vials were sealed with parafilm to prevent evaporation and then stored at 4˚C for 48 h. The concentrations of Chl a, Chl b and Cx + c were measured using a spectrophotometer at 662, 644 and 470 nm respectively. A solution of 95.5% acetone was used as a blank. The Chl a, Chl b, total chlorophyll and carotenoid concentrations in the leaf tissues were calculated according to the following equations and expressed as mg・g−1 fresh mass:
Chl a = 9.784D662 − 0.99D644.
Chl b = 21.42D644 − 4.65D662.
Total chlorophyll = Chl a + Chl b.
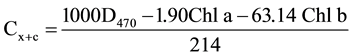
where Di is an optical density at the wavelength i.
3. Statistical Analysis
Each experiment was repeated thrice with each treatment sample containing ten individual plants and data presented are mean ± standard error (SE). The results were subjected to one way ANOVA and LSD test was used for comparison between pairs of treatments. The data analysis was carried out using MS excel 2003 and statistical package, SPSS 10.
4. Results and Discussion
4.1. Morphological Changes in Response to Arsenic and Curcumin Treatment
The effect of arsenic on germination and growth of mung bean seedlings was in accordance with the earlier report [25] . Arsenic induced morphological, physiological, biochemical & an anatomical change in plants is well known [25] . However, physiological, biochemical and anatomical changes occurs at the early stage of toxicity than the morphological changes. In the present study, arsenic induced morphological changes were least visible, but physiological and growth analysis showed growth reduction in mung bean seedling. Some of the plants under arsenic stress showed constriction of leaves with appearance of burning spots in the leaf apex. However, at the lower concentration of curcumin (25 μM) better growth and ameliorating effect was evident by healthy growth of the mung bean seedlings when supplied alone or in combination with arsenic. Such effect of curcumin may be attributed to its antioxidant potential which protects cell from arsenic induced oxidative damage in mung bean. However, there is no report on the biological role of curcumin in plants.
4.2. Effect of Arsenic and Its Interaction with Curcumin on Seed Germination
The inhibitory effect of arsenic on the germination of mung seeds was evident from relatively lower germination rate (28%) in arsenic treated seeds as compared to control (84%) (Table 1). The germination percentage of curcumin treated seed was found to be 75%, 15%, 10% at 25, 50, & 100 μM treatment respectively. Seed germination percentage reduced under arsenic stress but curcumin at 25 μM concentration showed protective effect on seed germination when treated alone or in combination with arsenic. The length of radicle growth also varied among the treatments as shown in Figure 1.
4.3. Arsenic Induced Changes in Growth of Mung Bean Seedling
An exposure to As (50 μM) caused significant reduction in root and shoot length of mung bean seedlings (Figure 2). This inhibitory effect was more pronounced on root length than shoot length. The root length decreased by over 49.3% in response to 50 μM As. In contrast the reduction in shoot length was comparatively lesser (15.13%) which was also evident from declining root-shoot ratio upon As treatment (as depicted in Figure 3(B)). Further as compared to control root dry mass decreased by 75% whereas no significant changes in shoot dry
![]()
Table 1. Effect of arsenic, curcumin and their interaction on Chl a, Chl b, Chl a:b, total Chlorophyll, total Carotenoid contents of growing seedings & seed germination of mung bean [Vigna radiata (L.) Wilczek]. Data presented are mean ± SE (n = 3). Mean value superscript with “a” & “b” indicates significant mean difference from control & arsenic treatment at p = 0.05 in multiple comparison test by LSD respectively.
![]()
Figure 1. Effect of As and its interaction with Curcumin on germination of mung [Vigna radiata (L.) Wilczek] seeds.
mass was observed (Figure 4). However, the total dry mass of mung bean seedlings decreased by 24% in As treated seedlings as compared control. Azad et al. [26] also reported a decline in shoot length in Oryza sativa at higher concentration of arsenic. Arsenate in plants is absorbed and translocated through the phosphate channels [27] [28] . In our present study, the root growth was significantly inhibited than shoot growth. This may be attributed to the poor translocation of arsenate to the shoot due to the antagonistic effect of phosphate ions present in the nutrient solution on are senate for root to shoot translocation through the phosphate channels or the increased level of chelation of arsenate in the roots.
4.4. Arsenic Induced Changes in Photosynthetic Pigments
The results show that As also induces changes in photosynthetic pigment. In comparison to control, chl a, chl b, total chlorophyll and carotenoid content decrease by 68.25%, 69.98% and 27.03% respectively in shoot of
![]()
Figure 2. Effect of arsenic (50 μM), curcumin (25, 50 & 100 μM) and their interaction [50 μM As + curcumin (25, 50 & 100 μM)] on root (A) and shoot (B) length of growing seedlings of mung bean [Vigna radiata (L.) Wilczek]. Data presented are mean ± SE (n = 3). Mean value superscript with “a” & “b” indicates significant mean difference from control & arsenic treatment at p = 0.05 in multiple comparison test by LSD respectively.
![]()
Figure 3. Effect of arsenic (50 μM), curcumin (25, 50 & 100 μM) and their interaction [50 μM As + curcumin (25, 50 & 100 μM)] on root (A) and shoot (B) dry mass of growing seedlings of mung bean [Vigna radiata (L.) Wilczek]. Data presented are mean ± SE (n = 3). Mean value superscript with “a” & “b” indicates significant mean difference from control & arsenic treatment at p = 0.05 in multiple comparison test by LSD respectively.
![]()
Figure 4. Effect of arsenic (50 μM), curcumin (25, 50 & 100 μM) and their interaction [50 μM As + curcumin (25, 50 & 100 μM)] on root (A) and shoot (B) dry mass of growing seedlings of mung bean [Vigna radiata (L.) Wilczek]. Data presented are mean ± SE (n = 3). Mean value superscript with “a” & “b” indicates significant mean difference from control & arsenic treatment at p = 0.05 in multiple comparison test by LSD respectively.
growing mung seedlings. Chl a: Chl b ratio increased by 5.75% in arsenic treated mung seedlings. Srivastava et al. [29] also reported a similar decline of chl a, chl b content in Hydrilla verticillata at higher doses of arsenic treatment. This decrease in chlorophyll synthesis could be due to the generation of reactive oxygen species like superoxide and hydroxyl radicals, and hydrogen peroxide that have the potential to damage proteins, nucleic acids and amino acids involved in the biosynthetic pathway of chlorophyll synthesis [30] .
4.5. Effect of Curcumin and Its Interaction with As on Growth of Mung Bean Seeds
Curcumin causes significant changes in growth of mung seedlings. Although changes in root length of mung bean is lower relative to control but compare to arsenic treated seedlings curcumin treated samples showed increase in root length. It was evident that As induced decrease in root growth is checked when curcumin and arsenic is treated simultaneously. There is no significant changes in shoot length in curcumin treated plants relative to control. But in comparison to arsenic treated sample, increase in shoot length due to curcumin treatment was observed. The interaction of curcumin with As however could not increase any shoot length. The root dry mass was decrease with curcumin treatment, but relative to control root dry mass shows no significant changes when 50 μM As and 25 μM curcumin are interacted. It also clearly indicates that the toxic effect of arsenic in root of growing mung seedlings is minimized by increasing root biomass over 350% when the plant is supplied with 50 μM As along with 25 μM curcumin. Although As have no significant effect on shoot biomass, curcumin increases shoot biomass by 11.64% at 50 μM concentration relative to control. The interactive effect of arsenic with curcumin also showed increase in shoot biomass over 23.52%, 17.64% and 11.76% in arsenic treated plant at 25, 50, 100 μM curcumin interaction respectively. Shoot dry mass increased because of curcumin interaction by 23.52%, 17.64%, 11.76% at 25, 50, 100 μM curcumin concentration respectively with respect to arsenic treated plant. However the total dry mass increases by 20% in plants treated with arsenic and curcumin (50 μM As + 25 μM curcumin) relative to control. It also clearly indicates that total dry mass has increased by 57.89% due to curcumin interaction (50 μM As + 25 μM curcumin) in comparison with As treated plants. This findings highlight the fact that curcumin prevents cell biomembranes damage. Lipid peroxidation is a chain reaction triggered by the free radicals produced in plants under stress conditions, leading to the damage of the cell membranes. Curcumin checks lipid peroxidation by scavenging the reactive free radicals that mediate the process of lipid peroxidation [31] [32] . Curcumin exhibits the free radical scavenging activity by donating H-atom from the phenolic group in acidic or neutral pH [33] [34] .
4.6. Effect of Curcumin and Its Interaction with As on Photosynthetic Pigments
The interaction of curcumin with As shows changes in photosynthetic pigments. In comparison to control, chl a, chl b, total chlorophyll and carotenoid content decreased in shoot of growing mung seedlings as already mentioned. However in comparison with As treated plant, curcumin caused increase in chl a, chl b and apparently, total chlorophyll increased by 175.91%, 186.11%, 178.69%, 230.11%, 42.86% & 26.77% in mung bean shoot at 25, 50 & 100 μM curcumin and its interaction with As (50 μM As + 25 μM curcumin; 50 μM As + 50 μM curcumin & 50 μM As + 100 μM curcumin) treatment respectively. The effect of curcumin and its interaction indicates that there was a decline in chlorophyll ratio highest being shown at 50 μM As + 25 μM curcumin treated mung seedlings. Total carotenoid content also increased due to curcumin interaction highest being shown by several thousand fold increase in mung seedlings treated with As & curcumin (50 μM As + 50 μM curcumin & 50 μM As + 100 μM curcumin) respectively relative to only As treated plants. This increase in the total chlorophyll content can be attributed to the ability of curcumin to interact with DNA and mediate the transient increase in expression of enzymes of the C5 pathway for chlorophyll synthesis [35] . Such effect of curcumin may be due to its potentiality to act as antioxidant as reported elsewhere in animal cells [36] - [39] .
The effect of arsenic on germination and growth of growing mung bean [Vigna radiata (L.) Wilezek] is found to be inhibitory, which is in consistent with earlier reports in mung bean [25] and in other plants. In the present study, reduction in root elongation was accompanied by decrease in shoot dry mass resulting into reduced growth in growing mung bean. Such reduction in growth may be due to anatomical changes in mung bean root as reported earlier [25] . We observe germination inhibition by arsenic in mung bean as well as the reduced growth in the growing seedlings. The inhibitory effect of arsenic was more pronounced in root than in shoot. Although, curcumin treatment did not induce seed germination at the level of the control plant when treated at different concentration (25, 50 & 100 μM), the interactive effect of curcumin was interesting. The seed germination in curcumin and arsenic treated seed showed 56%, 37%, & 38% relative to only arsenic treated seeds only with 28% at 25, 50, & 100 μM curcumin. It clearly indicates that curcumin at lower concentration (25 μM) can enhance germination in mung bean under arsenic stress. However, curcumin alone did not show significant increase in germination relative to control seed. Arsenic affects the plant by lowering the photosynthetic pigment content as reported in case of other stresses [40] -[45] . Pigment content and photosynthesis in various type of plant are generally reduced by arsenic and other metal stress. Chlorophyll a & b decreased in the leaves of mung bean due to arsenic stresses, consequently lowering the total chlorophyll. Such decrease in chlorophyll many enhance chlorosis & necrosis in plant as reported in case of other abiotic stresses. The reduction in carotenoid content due to arsenic stress may increase photosynthetic damage. However, total carotenoid is relatively higher than total chlorophyll which more prominently increased due to curcumin and its interaction with arsenic. Such increased carotenoid content may be an adaptive response to cope with stress induced photosynthetic damage.
5. Conclusion
From the present study, it may be suggested that the arsenic induced growth reduction in mung bean seedlings with simultaneous reduction in photosynthetic pigments but the curcumin, alleviates the adverse effect of arsenic on mung bean seedlings. It can also be suggested the arsenic induced germination inhibition of mung bean may be counteracted by curcumin at lower concentration. Further detail study on the curcumin effect on physiological events during mung seed germination and physiochemical and antioxidative responses of growing seedlings of mung bean (Vigna radiata) can reveal the exact mechanism underlying the amelioration of arsenic toxicity by curcumin in growing seedlings of mung bean (Vigna radiata).
Acknowledgements
Authors gratefully acknowledge the Department of Biotechnology, Govt of India, for providing research support under the Biotech Hub (San. No.: BT/04/NE/2009 dated 21/09/2010) and Star College programs (San. No.: BT/ HRD/11/04/2011 dated 21/03/2011) to Karimganj College, Karimganj, Assam, India.