Ultrastructural Identification of the Basal-Granulated Cells in the Duodenum of Albino Rat ()
1. Introduction
The origin and differentiation of different types of basal-granulated cells in the gastro-enteric mucosa of man and animals are subjects gaining much popularity in laboratories nowadays. By the use of the Wiesbaden terminology these cells had been identified as enterochromaffin (EC) cells that produce biogenic amines and/or polypeptide hormones [1] . The occurrence of such endocrine cells has long been known by many authors [2] -[10] . As far as their fine structure, the electron microscope has helped in differentiation of their types in tissues taken from the stomach, duodenum, colon and rectum in man [2] -[7] . Immunocytochemical studies had also been done to determine the nature of the hormonal peptides liberated by each cell type [11] -[14] . It now seems desirable to collect, in addition, morphological data on various species of animals to find the fine cellular structure and the possible source of hormones of such highly specialized cells. As far as the albino rat is concerned, the ultrastructural differentiation of the basal-granulated cells in the duodenum is lacking.
In health, enteroendocrine cells (EEC) play multiple roles in the gastrointestinal response to food, coordinating secretory and motor events to maximise the efficient digestion and absorption of food. They also play a key function in maintaining epithelial integrity, and contribute to the mucosal innate immune system. An important further role is the short-term control of food intake [15] .
These cells are distributed in the stomach and intestines of vertebrates, 1% of which are present in the intestinal epithelia and represent the largest population of hormone-producing cells in the body [16] and [17] . They control the secretion, absorption, motility, and proliferation of epithelial cells [18] and [19] .
The neuroendocrine cells, dispersed among the epithelial cells of the gastrointestinal tract, together with the enteric nervous system play a vital role in the function of the digestive system. Great interest in the role of these endocrine cells and the enteric nervous system increased following the demonstration of their involvement as neurotransmitters in the regulation of muscular movement, secretion of intestinal glands and control of vascular permeability of the digestive system [20] .
Immunohistochemical studies have been performed to investigate the distribution and relative frequency of neurons and neuroendocrine cells containing these neurotransmitters and neuropeptides in the gastrointestinal tract of several mammals including humans [21] , horse [22] , pigs [23] , lesser mouse deer [24] and rat [25] in order to elucidate their function.
The aim of the present study was, therefore, to represent the distribution profile as well as the fine structure of the basal granulated cells in albino rat duodenum.
2. Material and Methods
The material was obtained from twenty healthy albino rats free of any obvious disease. All animals were males with an average weight of 350g. Anaesthesia was made by intraperitoneal injection of 3% solution of chloralhydrate; 1 ml/l00g body weight. Effective perfusion was done by 3% glutaraldehyde in 0.2 M cacodylate buffer at pH 7.4 for 10 minutes [26] . After perfusion, samples of the duodenal mucosa were cut perpendicular to the long axis into 1 mm3 pieces, fixed further by immersion for 3 hours in 4% glutaraldehyde and rinsed with sucrosecacodylate buffer. The specimens were postfixed in 1% osmium tetroxide, dehydrated and embedded in epon. Ultrathin sections were prepared, stained with uranyl acetate and lead citrate [27] and examined by a transmission electron microscope.
3. Results
In all of the specimens examined, cells with secretory granules were dispersed mainly in the epithelium of the crypts of Lieberkuhn (Figures 1-7).
The granules were of variable shapes and sizes and accumulated usually in the basal parts of cytoplasm. The fine structure of the different cells revealed some common features. All cells were in direct contact with the basal membranes of the crypts. The nucleus was generally oval. Mitochondria, free ribosomes, rough endoplasmic reticulum and lysosomes were observed in their cytoplasm (Figures 1-7).
The outline of such basal-granulated cells was variable from one type to another but generally they were characterized by the presence of electron-dense granules. Differentiation of several types was done on the basis of the morphology of the granules.
Cell type I: were enterochromaffin cells (EC) as revealed in the electron micrographs by the shape and size of their secretory granules (Figure l). They constituted the majority of the cells in question. The granules were concentrated between the nucleus and basal lamina. All of the basal granules were heavily contrasted and of characteristically rod-like, ovoid, tortuous or kidney shaped (Figure 2).
Cell type II: were EC-like cells. The granules of this cell type resembled those of type I in shape and electron density but were seen in the whole cytoplasm (Figure 3).
Cell type III: was characterized by medium-sized round granules which vary in electron density (Figure 4).

Figure 1. An electron micrograph of albino rat duodenal crypt epithelium showing a part of an enterochromaffin (EC) cell with dense irregular granules between the nucleus (N) and basal lamina (B). The EC-cell does not reach the lumen (L). (Mic. Mag. ×2600)

Figure 2. A high magnification of a part of an enterochromaffin (EC) (arrow) cell from albino rat duodenal epithelium. Note the characteristic rod like or tourtous dense basal granules (arrows). N = nucleus M = mitochondria. (Mic. Mag. ×10,000)
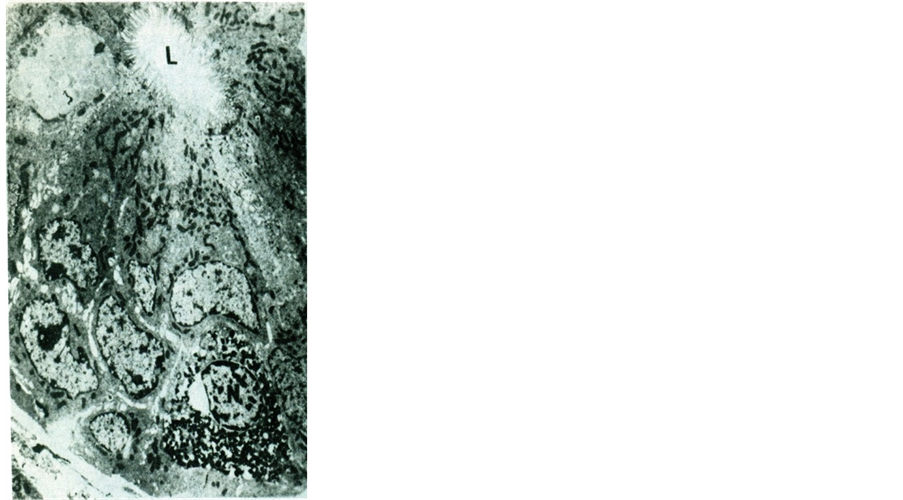
Figure 3. An electron micrograph of albino rat duodenal crypt epithelium showing a large number of irregular heavily concentrated secretory granules seen in the whole cytoplasm. Note that the cell does not reach the lumen (L) of the crypt. N = nucleus. (Mic. Mag. ×2600)

Figure 4. An electron micrograph of albino rat duodenal crypt epithelium showing another basal granulated cell (M) with medium even sized round granules varing in electron density (arrow). N = nucleus, ER = rough endoplasmic reticulum. (Mic. Mag. ×4000)
Cell type IV: showed large round granules with homogenous substance. The electron density of the specific granules was lower than in cell type 1 (Figure 5).
Cell type V: was characterized by small round granules of lower electron density. This type of cells was slender in shape. The granules were small and in contact with the basement membrane in a small area. Small mitochondria were seen separating the granules (Figure 6).
Cell type VI: This type possessed few granules of small size and solid appearance. Large electron lucent granules were also intermingled between the smaller solid ones (Figure 7).
4. Discussion
The occurrence of different types of basal-granulated or endocrine cells in the gastro-enteric mucosa of man and animals raised the need to study the fine structure of such cells in the albino rats. The morphological features of these cells are variable from one species to another, but generally they are characterized ultrastructurally by the presence of secretory granules of variable shape and size. The present study reported the occurrence of six different types of endocrine cells in the albino rat duodenal mucosa. The fine structure of each type had been described. All of the cells in question showed variable kinds of different sizable granules of varying electron density.
Entero-chromaffin (EC) cell was the predominant cell type, with its characteristic rod-like or tortuous granules. A second cell type with granules that resembled those of the EC but were seen in the whole cytoplasm (EC-Ii e cells) was also described. M-cells whose granules appeared medium and even-sized round ones were identified as a third type. A fourth type showed large, round and dense granules (L-cells). S-cells showed their granules in contact with the basement membrane in a small area and small mitochondria separating them. A sixth type with large electron lucent granules intermingled between smaller solid ones was also identified.

Figure 5. An electron micrograph of albino rat duodenal crypt epithelium showing basal granulated cells (L) with large round granules of homogenous substance (arrow). Note again the cell does not reach the lumen (Lu) of the crypt. N = nucleus. (Mic. Mag. ×4000)

Figure 6. An electron micrograph of albino rat duodenal crypt epithelium showing a basal granulated cells (S) with small granules in a small area and in contact with the basement membrane (B). Small mitochondria are seen. Note that the S-cell is covered by the surrounding crypt epithelial cells. (Mic. Mag. ×5000)

Figure 7. An electron micrograph of albino rat crypt epithelium showing basal granulated cells (G) with different sizable granules varying in electron density. The large granules are electron lucent and the small ones are dense and solid. P = Paneth cell. Note that the part of the G-cell is seen in the field does not reach the lumen (L). (Mic. Mag. ×5000)
Neuroendocrine cells of the gut are highly specialized mucosal cells that produce a wide range of hormones with specific regional distribution [18] .
It should be taken into account that, in gastric and intestinal tissues of man and all animal species so far examined in previous studies demonstrated a similar fine structure of such endocrine cells as was reported in the present study [2] -[10] . But, in contrast to the description given to such cells as open type endocrine cells found in other animal species as dog and rabbit [5] [8] , those basal-granulated cells in the duodenal mucosa of albino rats were all of the close type cells. All of the cells described in the present study were closed in type as they were isolated from the lumen of the crypts by other cells covering them. The cells which are closed in type might receive mechanical stimuli such as pressure and tension caused by meals and passing faeces. While in the opened type endocrine cells, a different perceptional mechanism postulated that such cells are a kind of chemoreceptor which might perceive chemical and physiological information from the lumen to control its own secretory activity [28] [29] .
The heterogeneity and concentration of neuroendocrine cells in the duodenum may be related to regulation of the secretion of pancreatic juice and bile as well as to the functional control of the small intestine [30] [31] . In fact many neurotransmitters and neuropeptides have been identified in the pancreas of the one-humped camel [32] .
Differences in the distribution pattern of endocrine cells in the intestinal segments are related to their physiological functions in region or intestinal segments. According to Reference [33] , the endocrine cells are mainly involved in controlling the release of digestive enzymes in the small intestine and in controlling intestinal motility in the large intestine.
In accordance with the present findings, the ultrastructural differentiation of such a group of basal-granulated cells in the duodenal mucosa of albino rats might enable the physiologists, internists and other research workers to study several biologically active peptides with well established functions other than those which have long been studied [11] -[14] . Correlation between the distribution of such cells in the intestine of albino rats and others in the central nervous system of the same animal awaits also classification.
Acknowledgements
I would like to express my very great appreciation to Professor M. Shaaban Farag. Professor and Head of department of Anatomy & Histology, Salman bin Abdulaziz University, Al Kharj, KSA. I am particularly grateful for the assistance given by him. He provided me with very valuable experience and enthusiastic encouragement throughout the research.