Mathematical Analysis of a Large Scale Vector SIS Malaria Model in a Patchy Environment ()
1. Introduction
We recall the large scale system developed in [1] reduced into a compact form as
 (1)
(1)
where
 , is a vector representing;
, is a vector representing; 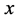 is the proportion of infectious children,
is the proportion of infectious children, 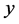 is the proportion of infectious adults, and
is the proportion of infectious adults, and  is the proportion of infectious mosquitoes.
is the proportion of infectious mosquitoes.
 ,
,  and the matrix
and the matrix  are the matrices.
are the matrices.


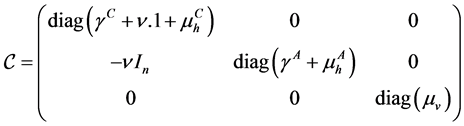
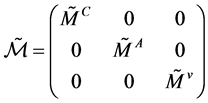
The authors used the preceding matrices and the vector 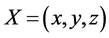 to rewrite Equation (10) in [1] in a compact form as
to rewrite Equation (10) in [1] in a compact form as
 (2)
(2)
This system evolves on the unit cube of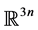 .
.
Calculation of the Basic Reproduction Number
We use the classical framework defined in [3] [4] .
The application  represents the rate of appearance of new infections in the compartments in the patches.
represents the rate of appearance of new infections in the compartments in the patches.
The function  is the rate of transfer of individuals in compartments.
is the rate of transfer of individuals in compartments.
If  is set to zero, system becomes
is set to zero, system becomes , which is a linear system, and we have already seen that
, which is a linear system, and we have already seen that  is a stable Metzler matrix.
is a stable Metzler matrix.
Proposition 1.1
The basic reproduction number of system (1) is
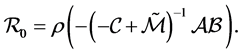
spProof
This is straightforward since the Jacobian of  computed at the DFE
computed at the DFE  is
is

and the Jacobian of  computed at the DFE is
computed at the DFE is .
.
The next generation matrix is then  □
□
We can develop the expression of  further:
further:
 (3)
(3)
 (4)
(4)
Then we can compute the nonnegative matrix , which is a lower triangular matrix
, which is a lower triangular matrix

with
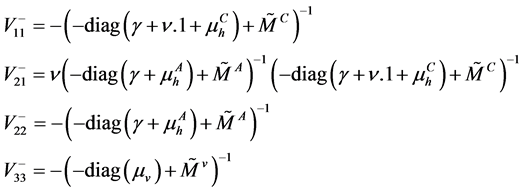
The next generation matrix is a block matrix
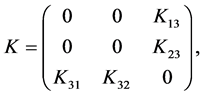
with
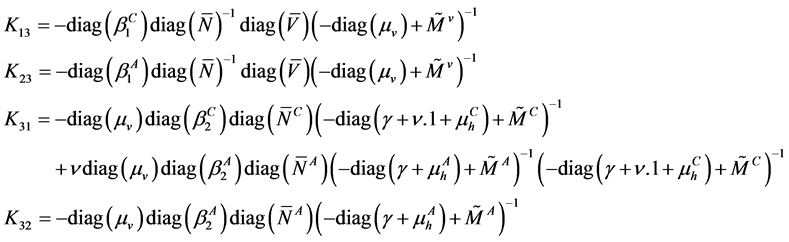
The block structure of  implies (see [4] ) that
implies (see [4] ) that

When , the DFE is locally asymptotically stable, and if
, the DFE is locally asymptotically stable, and if 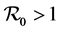 the DFE is unstable, see [3] [4] .
the DFE is unstable, see [3] [4] .
2. Main Result
In this section we establish a global stability result for the DFE when 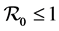 and a global stability result when
and a global stability result when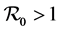 . We have the following theorem
. We have the following theorem
Theorem 2.1
We consider the system (1) with the matrix 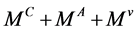 irreducible.
irreducible.
Then If , then the system (1) is globally asymptotically stable at the origin If
, then the system (1) is globally asymptotically stable at the origin If , then there exists a unique endemic equilibrium
, then there exists a unique endemic equilibrium , which is globally asymptotically stable on
, which is globally asymptotically stable on 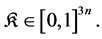
spProof
We recall system (1),

The Jacobian at the origin will be given by

and
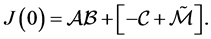
To prove the first part of the proposition above we assume that  Following [5] ,
Following [5] , 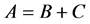 is a regular splitting of A if B is Metzler stable and
is a regular splitting of A if B is Metzler stable and . Thus in our case
. Thus in our case 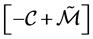 has to be a stable Metzler matrix which is invertible and
has to be a stable Metzler matrix which is invertible and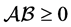 , or equivalently,
, or equivalently, 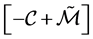 has to be an M-matrix.
has to be an M-matrix.
We know from Thieme [6] , Driessche [4] , and Varga [7] that

From the preceding section we know that the Jacobian  is an irreducible Metzler matrix. So, by PerronFrobenius, there exists a positive vector
is an irreducible Metzler matrix. So, by PerronFrobenius, there exists a positive vector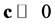 , such that
, such that

To prove the global stability of the DFE we consider the Lyapunov function
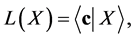
where  denotes the inner product. From the definition of
denotes the inner product. From the definition of , this function is actually positive definite in the nonnegative orthant.
, this function is actually positive definite in the nonnegative orthant.
We compute the derivative of 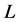 along the trajectories of (1) and find that it is equivalent to
along the trajectories of (1) and find that it is equivalent to
 (5)
(5)
We see that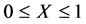 , it is clear that
, it is clear that , hence the above inequality.
, hence the above inequality.
Since  the derivative is non positive. The DFE is stable.
the derivative is non positive. The DFE is stable.
We will prove the asymptotic stability when 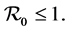
First we consider the case when . Since we know that
. Since we know that  implies
implies ,
,  is negative definite, since
is negative definite, since . This proves the asymptotic stability of the DFE.
. This proves the asymptotic stability of the DFE.
When , we consider the largest invariant set contained in the set
, we consider the largest invariant set contained in the set
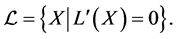
For such an  we have
we have

but since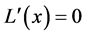 , we have by the inequality (5),
, we have by the inequality (5),

Hence

Since  then
then 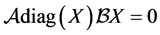 or equivalently
or equivalently 
Now we show that the largest invariant set in 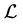 is reduced to the origin.
is reduced to the origin.

We must have, for any index ,
, 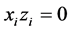 and
and 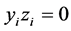
Suppose  since
since

we have

Then 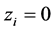 and
and 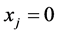 for any patch
for any patch , with a “children” arc leaving
, with a “children” arc leaving  and entering
and entering .
.
Since  and
and  and since
and since
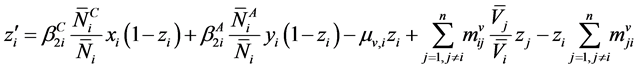
we have

Again  and
and 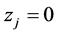 for any patch
for any patch 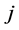 with a “mosquito” arc leaving
with a “mosquito” arc leaving  and entering
and entering .
.
Now 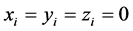 implies
implies
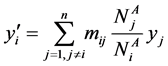
which implies that 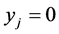 for any patch with a “adult” arc leaving
for any patch with a “adult” arc leaving  and entering
and entering .
.
Now, since any patch can be reached by a path composed of “children”, “adult” or “mosquito” arcs, this proves that  for any index.
for any index.
This ends the proof for the global asymptotic stability of the DFE from LaSalle’s Invariance Principle [8] .
To prove the second part of our theorem, when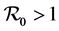 , we need the following theorem from [9] .
, we need the following theorem from [9] .
Theorem 2.2
Let  be a
be a  vector field in
vector field in , whose flow
, whose flow  preserves
preserves  for
for  and is strongly monotone in
and is strongly monotone in . Assume that the origin is an equilibrium and that all trajectories in
. Assume that the origin is an equilibrium and that all trajectories in  are bounded. Suppose the matrixvalued map
are bounded. Suppose the matrixvalued map 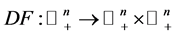 is strictly anti monotone, in the sense that,
is strictly anti monotone, in the sense that,
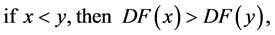
then either all the trajectories in  tend to the origin, or else there is a unique equilibrium
tend to the origin, or else there is a unique equilibrium  and all the trajectories in
and all the trajectories in  tend to
tend to .
.
For our case we shall consider the positively invariant set , which is diffeomorphic to the nonnegative orthant
, which is diffeomorphic to the nonnegative orthant . Since the faces of the cube of type
. Since the faces of the cube of type  are repulsive for the vector field associated to (1), all the trajectories are bounded in
are repulsive for the vector field associated to (1), all the trajectories are bounded in .
.
We recall system 1.
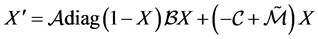
If we take , then
, then

Clearly
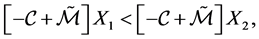
since the quantities are positive.
Now we prove that
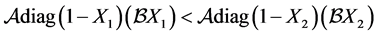
and hence show that the system is strongly monotone. That is

or

Considering the structure of 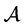 and
and  and having
and having  in
in 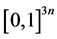 and the fact that a sign change reverses the inequality, then
and the fact that a sign change reverses the inequality, then

hence

To prove that theorem, we recall the Jacobian  of system (1)
of system (1)

Again for any 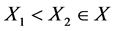 , then
, then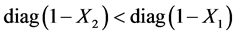 . Since the matrix
. Since the matrix  has on each row a positive term, since
has on each row a positive term, since  is a diagonal matrix with positive terms, we deduce
is a diagonal matrix with positive terms, we deduce
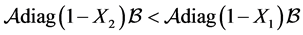
Considering the structure of  we have, if
we have, if , the relation
, the relation 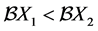 holds and consequently
holds and consequently 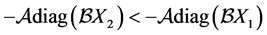 .
.
Finally we have , therefore the anti monotone criteria is met.
, therefore the anti monotone criteria is met.
We will prove that no trajectory tends to the origin.
We have 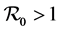 which is equivalent to
which is equivalent to . Then there exists a positive vector
. Then there exists a positive vector  such that
such that

We consider the Chetaev function on a neighborhood of the origin

An simple computation gives
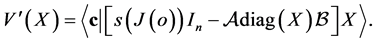
Then in a sufficiently small neighborhood of the origin, in ,
, . This proves that for
. This proves that for 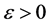 sufficiently small, the hyperplane
sufficiently small, the hyperplane  is a barrier for the vector field associated to (1). This proves that no trajectory tends to the origin. Then we conclude, by Hirsch theorem, the existence of an attractive endemic equilibrium
is a barrier for the vector field associated to (1). This proves that no trajectory tends to the origin. Then we conclude, by Hirsch theorem, the existence of an attractive endemic equilibrium 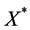 in the interior of the cube.
in the interior of the cube.
To prove stability, we shall compute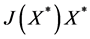 :
:
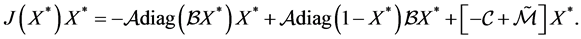
Taking into account that 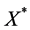 is an equilibrium gives
is an equilibrium gives
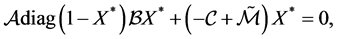
therefore

since 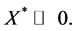
We have proved that there exists a vector  such that for the Metzler matrix
such that for the Metzler matrix  we have
we have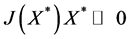 . This implies that
. This implies that  is Hurwitz [10] [11] .
is Hurwitz [10] [11] .
This completes the proof of the global asymptotic stability of the endemic equilibrium.
3. An Example in Two Patches
In this section we give a result to the case of two patches. We shall use the structure defined in Subsection 1.1.

The basic reproduction number is given by 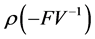 From our example and at the DFE, this matrix is defined by
From our example and at the DFE, this matrix is defined by 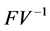
which has the values
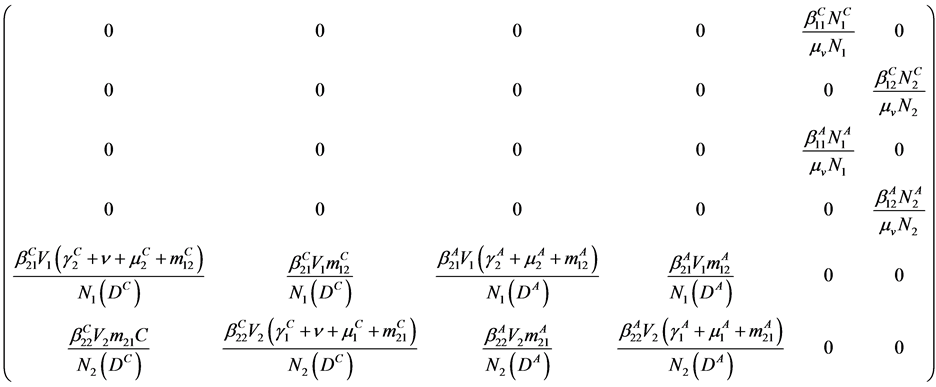
where
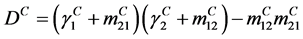 ,
, . To get the basic reproduction number we need to solve
. To get the basic reproduction number we need to solve  which is a
which is a  matrix. Rewriting the matrix in the form
matrix. Rewriting the matrix in the form

The determinant of 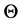 is given by
is given by

Using the properties of determinants we have
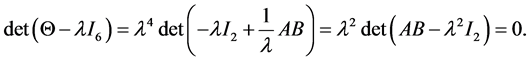
We see after some calculation, that

where
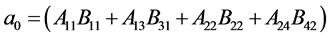

The expression for , is complex due to the large number of parameters involved, but from the expression of the matrices A and B above, we can gain some insight. For example if there is no human migration between the two patches, then
, is complex due to the large number of parameters involved, but from the expression of the matrices A and B above, we can gain some insight. For example if there is no human migration between the two patches, then 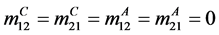 and
and
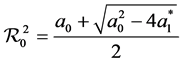
where

This is the product of the maximum basic reproduction number for patch 1 and patch 2.
If there is no infective vectors in patch 2, and no vector migration, then , and
, and

which is the total children and adult contribution to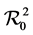 .
.
It is clear that this new value of the basic reproduction number highly depends on the migration rates of the two age groups. If we increase the migration rates then  increases.
increases.
Biologically, this implies that back and forth movement between the patches, would introduce malaria infection in an otherwise malaria free patch.
4. Simulation
In this section we obtain baseline values for two sites: the U-shaped valleys and V-shaped valleys. and use them to simulate equation 1, which is a compact form of equation 5 in [1] . For the human population in our model, we consider two patches, Umutete and Iguhu for the U-shaped valleys, and, Marani and Fort Tenan for the V-shaped valleys. From the study on the different ecosystems, the plateaus and the U-shaped valleys ecosystem have the characteristic, such that the results for the V-shaped valleys apply to the plateau ecosystem. This results from the fact that on the plateaus, the terraine is characterised by raised but flat topography with very little stagnant water as the water darains down the rivers, to support breeding places for mosquitoes. The only notable differences is where there are large water bodies like dams and reservoirs. In these cases high mosquito population is likely to survive and hence increase malaria transmission and infection. Some suitable references for our values are [12] -[16] .
Data for , and
, and  was estimated to be 10000 people and we assume that the population is evenly distibuted to the two patches so that we have 5000 people on each patch (1500 children and 3500 adults for each ecosystem). The mosquito population likewise was estimated to be 80,000 mosquitoes in the U-shaped valleys and 10,000 mosquitoes in the V-shaped valleys.
was estimated to be 10000 people and we assume that the population is evenly distibuted to the two patches so that we have 5000 people on each patch (1500 children and 3500 adults for each ecosystem). The mosquito population likewise was estimated to be 80,000 mosquitoes in the U-shaped valleys and 10,000 mosquitoes in the V-shaped valleys.
The summary of the parameter values used is given in Table1
4.1. The U-Shaped Valley Sites: Iguhu and Umutete, When 
When the age structuring is considered, the dynamics of the host population in the U-shaped valleys is represented on Figure 1. When there is no age structuring, the dynamics for the U-shaped valleys are shown in Figure 2. If we consider the U-shaped and the V-shaped valleys as one epidemiological region representing Western Kenya, then the dynamics are represented by Figure 3. The disease in the age structured model fades out faster. The steady states also settle to the endemic equilibrium faster.
If there is no spatialization the values for the U-shaped valleys for both ecosystems has host population variation represented on Figure 3. The interaction between the patches raises infection rate, so that the disease persists in the total population, while it fades out fast when the patches are isolated.
4.2. The V-Shaped Valley Sites: Fort Tenan and Marani, 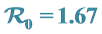
The dynamics of the model in the V-shaped valleys sites with age structure is given by Figure 4. When the age structuring is ignored the variation of the host population in the V-shaped valleys is represented by Figure 5.
5. Conclusions
Highland malaria in Western Kenya remains a source of mortality and morbidity. Concerned efforts have been put in place by the stakeholders to bring the disease under control with less than expected results. This study

Table 1 . Parameter values and ranges for System 1 and Equation (5) in [1] .
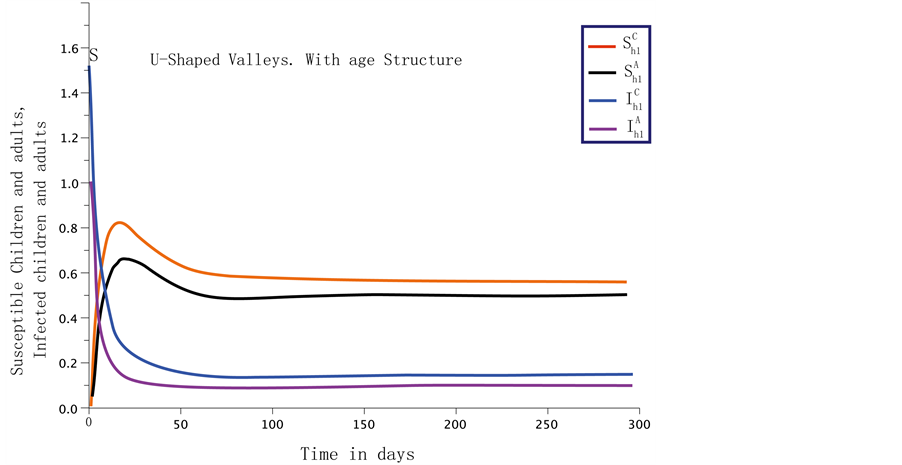
Figure 1. A numerical simulation for the variation of the two age classes population using Equation (1) and parameter values defined in Table 1 for the U-shaped valleys system with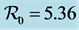 .
.

Figure 2. A numerical simulation of Equation (1) no age class in the population and .
.
captures important factors key to endemicity of of malaria in Western Kenya, which could direct control measure and targets effectively.
Age structure helps us differentiate between child’s infectivity and susceptibility to malaria infection. It is clear from Balls [17] that children are a significant source of mosquito infection compared to adults. The biting rate for the two age groups differs [18] [19] as children are bitten more than adults did. The other difference captured in the model is death rates for the children (which may include malaria induced deaths). Most malaria deaths occur in children under the age of five years. While the adults also suffer morbidity due to severe in-

Figure 3. The variation of the total population in the region, no age class and the two ecosystems are treated as a single U-shaped valley ecosystem.
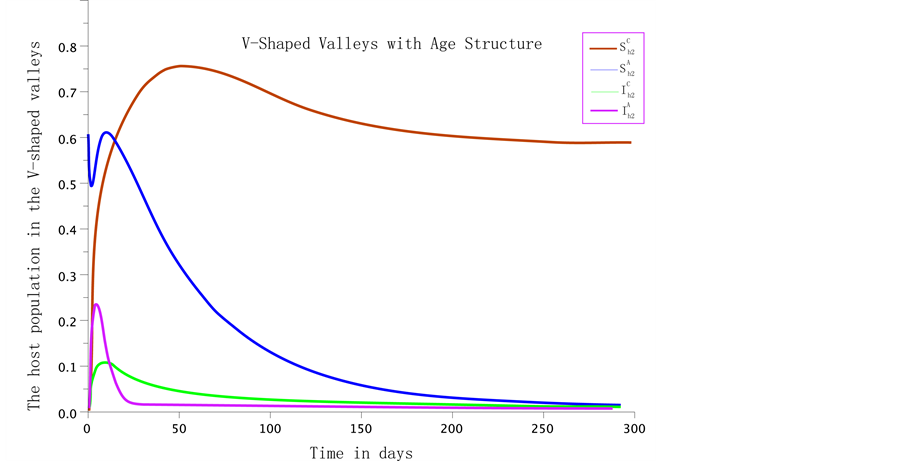
Figure 4. A numerical simulation of model 1 using parameter values defined in Table 1 for the V-shaped valleys ecosystem with age structure. In this case .
.
fection of highland malaria, there are fewer deaths due to acquired immunity compared to children. We note that the populations settle to the endemic equilibrium faster than in the age-structured than in the unstructured system as shown in Figure 5, and the stable equilibrium is achieved faster in the structured than the unstructured system. Adding age structure allows age specific control strategies to reduce disease prevalence.
Our model suggests that a suitable model for malaria should be one that captures: age structure; differentiated patch or region susceptibility, which depends on the immunity of the inhabiting population; differentiated infectivity, which also depends on the immunity and age of the host population, and the mosquito population
Figure 5. A numerical simulation of model 1 using parameter values defined in Table 1 for the V-shaped valleys ecosystem without age structure. In this case , A numerical simulation of model 1 using parameter values defined in Table 1 for the V-shaped valleys ecosystem. The is no age structures in the populations. In this case
, A numerical simulation of model 1 using parameter values defined in Table 1 for the V-shaped valleys ecosystem. The is no age structures in the populations. In this case .
.
dynamics. Intervention then can be done with guidance from the model.
A more comprehensive characterization of results would have to include other types of patches that may not be terraine related but have different epidemiological characteristics from the U-shaped and the V-shaped valleys. Such patches could take care of cities like Nairobi, where human migration has transferred malaria, and central Kenya where the cool highland ecosystem is disturbed by creation of dams for irrigation, rice cultivation, climate change and migration of population to the economically endowed part of the county. Adding age structure allows age specific control strategies to reduce disease prevalence.
We assumed that vectors migrate especially to nearby patches, and the migration parameters for hosts are constant, similar and independent of the compartment. For the compartments that are far apart, the migration of mosquitoes is negligible and is set to zero, since the mosquitoes are only able to fly about 2 kilometers away.
An explicit formula for 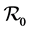 is obtained, which although complex due to the infinite number of patches, can be used to explore the effects of the parameters of the model. This formula will allow theoretical exploration of the options and efficiency of targeted public health intervention policies. The example in the two ecosystems simplifies the expression for
is obtained, which although complex due to the infinite number of patches, can be used to explore the effects of the parameters of the model. This formula will allow theoretical exploration of the options and efficiency of targeted public health intervention policies. The example in the two ecosystems simplifies the expression for , which we use to simulate our model with some realistic data from Western Kenya. This parameter is inversely related to migration of the hosts between the patches. This implies that to reduce
, which we use to simulate our model with some realistic data from Western Kenya. This parameter is inversely related to migration of the hosts between the patches. This implies that to reduce , we have to i) administer effective treatment through provision of proper health care facilities in both patches, ii) promote drug adherence, iii) reduce malaria drugs abuse through self administration to shorten the infectious period and arrest human to mosquito infection, hence increase the rate of recovery represented by
, we have to i) administer effective treatment through provision of proper health care facilities in both patches, ii) promote drug adherence, iii) reduce malaria drugs abuse through self administration to shorten the infectious period and arrest human to mosquito infection, hence increase the rate of recovery represented by .
.
An example in two patches is given with an expression of 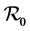 which is still complex; but insight is given in a case when no migration takes place. If the disease exists in one patch, with the back and forth movement, the disease in the otherwise free patch would be reintroduced. We want to mention that an example in three patches is also possible, but meaningful insight for the basic reproduction number may only be gained by simulation, with relevant data. This model can be extended to include intervention strategies by the Ministry of health in Kenya, through ITNs and IRS. Since the disease causes death especially in children, the model can also include disease related death rate in the human population. An important factor which is under investigation is the impact of climate related factors to the resurgent epidemics. Resistance of vectors to ITNs and IRS is also an important factor which may cause the disease to remain a menace in the region, not to mention the possibility of drugs resistance in human and possible emergence if new malaria strains.
which is still complex; but insight is given in a case when no migration takes place. If the disease exists in one patch, with the back and forth movement, the disease in the otherwise free patch would be reintroduced. We want to mention that an example in three patches is also possible, but meaningful insight for the basic reproduction number may only be gained by simulation, with relevant data. This model can be extended to include intervention strategies by the Ministry of health in Kenya, through ITNs and IRS. Since the disease causes death especially in children, the model can also include disease related death rate in the human population. An important factor which is under investigation is the impact of climate related factors to the resurgent epidemics. Resistance of vectors to ITNs and IRS is also an important factor which may cause the disease to remain a menace in the region, not to mention the possibility of drugs resistance in human and possible emergence if new malaria strains.
So far, we have formulated an analytical and numerical analysis which is a foundation of more research and also applicable to other vector borne disease like chikungunya [20] .
Acknowledgements
We wish to acknowledge of the Inria Metz, UMMISCO(IRD), the French Embassy in Nairobi and the university of Nairobi, Kenya, for their financial, logistic and moral, support during the writing of this article. We are very grateful to Dr. Githeko, KEMRI Kisumu for the great insight and literature he gave us during this study.
NOTES
*Corresponding author.