1. Introduction
Silver Oak (Brachylaena huillensis) is a versatile timber tree species in the family Asteraceae [1,2]. It is native to Central, East and Southern Africa. There has been a very high demand for B. huillensis wood and its products leading to over-exploitation. The tree species regenerate only through seeds and so far it is a threatened tree species [3,4]. The species is suitable for timber and carving artefacts [5] charcoal, essential oil, [5-7] sleepers, flooring blocks, furniture and turnery [5-7]. Moreover, due to its durability, the species is used as fence posts, building poles, transmission poles, ornamental and medicine for schistosomiasis and leaves are used for diabetes [6]. The Silver Oak is illegally exploited for timber, charcoal, carving, building poles, fencing posts, ornaments, medicine, perfumery and toilet preparations, sleepers, flooring blocks, furniture, and turnery [1,7,8]. B. huillensis regenerates through seeds. However, the seeds have poor germination rate. Also, B. huillensis seeds are difficult to collect because of their small size. Currently there is lack of a seed bank [5] and most of the seeds are eaten by insects, which renders the natural regeneration of the tree species uncertain.
In vitro micropropagation has proved in the recent as a means for supplying of planting material for forestry [9, 10]. In vitro plant culture offers advantages over conventional methods for multiplication and large-scale production of woody plants [11]. So far a good number of endangered and threatened species have been successfully regenerated using in vitro culture methods using shoot tips, leaves, and leaf bases [12]. Though, in vitro propagation in B. huillensis has not yet done and may be the best alternative method for propagating the tree species. Nevertheless, one of the major problems for many tissue culture systems is oxidized lethal browning and subsequent death of the cultured explants that usually depend on the phenolic compounds and the quality of the total phenols [13]. Phenolic compounds are secondary metabolites released from plants, which are present in high amounts. Browning in plants occurs mainly due to the oxidation of phenolic compounds by phenol oxidase [14]. This phenomenon occurs when the compartmentalized phenolic compounds are released during explant incision and henceforth react with phenolic oxidases and release quinone [14]. Quinone has negative effect on cell growth and can result in death/necrosis of cells [13]. Phennolic compounds occur as secondary metabolites in all plant species and they are generally characterized by a benzene ring and one hydroxyl group [14,15]. Plant phenolics are classified into major groupings distinguished by the number constitutive carbon atoms in conjuction with the structure of basic phenolic skeleton [15,16]. Many phenolics are rather reactive compounds and as long as no steric inhibition due to additional side chains occurs, they form hydrogen bonds [16].
The composition and synthesis of phenolics in plants tissue may be determined by genetic and environmental condition like oxidative reaction during culturing, processing and storage [17]. It appears that there is a relation between chemical compound of media and phenolic exudation, media discoloration, rooting deficiencies and explant browning and death. It was noticed that plant phenolic increased the rigidity of plant cell wall and acted as a molecular bridges between cell wall components [13]. During micropropagation, the exudation is very common and it often influences the results. Phenolic secretions and other exudates in plant tissue culture systems lessen explant initiation, growth, and development [18].
The antioxidant, ascorbic acid, was selected as it has been used successfully in the past to inhibit the exudation of phenols [19] and reduced oxidative browning in various plant species [20-22]. Ascorbic acid is able to scavenge oxygen radicals produced when the plant tissue is wounded, therefore protecting the cells from oxidative injury. The oxidative browning of explant tissue is reduced by ascorbic acid detoxifying these free radicals [23]. Moreover, ascorbic acid is an antioxidant that is able to prevent or inhibit oxidation process [24]. Besides its role as an antioxidant, ascorbic acid is involved in cell division and elongation [25]. Research by [26] on culture of banana cultivar Cavendish showed that ascorbic acid not only can prevent death due to explant browning, but also can increase the number of shoots growing on explant. The antioxidant compounds utilized in the experimental were selected because they have been used successfully in the past to delay browning in other woody monocotyledonous species. Thus, ascorbic acid is useful and effective in managing the problem of phenolics and improving plant growth in vitro [20]. However, excretion of phenolic compounds on in vitro culture can be controlled by various antioxidants in this study ascorbic acid was employed. Currently, in vitro propagation for B. huillensis via nodal segments is not yet in place, either a report regarding the effect of application of antioxidants in controlling lethal browning is lacking. In the present study the effect of ascorbic acid at various concentrations in controlling lethal browning using nodal segments of B. huillensis on culture was examined.
2. Materials and Methods
2.1. Plant Material and Explants Source
Plant materials were obtained from the healthy naturally growing trees in Bombo West Forest Reserve (BWFR). The BWFR is located in Korogwe district, Tanga, Tanzania. The reserve is owned by the central government; it was gazetted in 1959 with a Government Notice 1 of 1959 and has an area of 3523.5 hectares [27].
The tip branches of B. huillensis mature naturally growing trees were the source of explants material. Nodal segments of about ten centimeters were collected from the healthy naturally growing trees in Bombo West Forest Reserve in early April 2013. The nodes were preserved in a cool box with cold water and transported to Mikocheni Agriculture Research Institute (MARI) laboratory in Dar es Salaam. The nodes spent twenty four hours on transit before culture initiation.
2.2. Surface Sterilization of the Explants
In the laboratory, the nodes were placed in a bottle containing distilled water. The water contained two detergents, namely liquid soap and tween-20, which enhance the effectiveness of the disinfectant by breaking the surface tension between water and the plant tissues. For effectiveness, the nodes in the solution were agitated continuously for 10 minutes. Later the nodes were rinsed four times with distilled water. The bottles containing the already washed nodes were shifted to the transfer room and then immersed in sodium hypochlorite (NaOCl) 1.9% v/v) with two drops of tween-20 for 15 minutes. Later the nodes were rinsed four times with sterile distilled water and later dipped in 70% ethanol for 2 minutes. Thereafter, the nodes were rinsed four times using sterile distilled water before culturing. The sterilized nodes were trimmed properly to remove sterilizing agent affected parts. Nodal segments of about 1.5 centimeters with two or more nodes were cut from the sterilized nodes and cultured.
2.3. Culture Conditions and Media
Basal woody plant medium (WPM) [28] with full strength supplemented with 5 µM Benzylaminopurine (BAP) with four levels of ascorbic acid (0, 50, 100, 150, 200, & 250 mg/litre) was used. The pH of the medium was adjusted to 5.6 before autoclaving at 121 degrees centigrade. The surface sterilized explants (nodal segments) were inoculated on the WPM on the respective medium and labeled properly. Each magenta bottle with 40 ml of the WPM medium contained five explants. Three replications with 5 explants in each were maintained for each treatment and 15 explants in each treatment were evaluated. The culture free from ascorbic acid served as a control. The cultures containing the explants in magenta bottles were kept in a growth room at a temperature of 25 ± 2 degrees centigrade, 60% - 70% relative humidity and white fluorescent light with a 16-h photoperiod. Soon after initiation visual observation was made on daily basis, the number explants which browned were rated and recorded as follows; 1 = No discoloration, 2 = Slight discoloration, 3 = Moderate discoloration, 4 = Heavy discoloration, and 5 = extreme discoloration. The rating was modified from the rating scale given by [29].
3. Statistical Analysis
The obtained data were subjected to STATISTICA program and analyzed using one-way analysis of variance (ANOVA). The means are reported with standard errors. The fisher least significance difference (L.S.D.) was used to compare treatment means at p = 0.05 level of significance [30].
4. Results
The results in the present study showed that there was a significant difference among the concentrations of ascorbic acid used. However, there was no significant difference between ascorbic acid at 200 mg/l and 250 mg/litre concentration levels (Table 1). Generally it was revealed that there was a decrease in percentage of the extent of browning with an increase in concentration of ascorbic acid (Table 1). Relative to the control treatment, supplying ascorbic acid into the medium at the rate of 50, 100, 150, 200 and 250 mg/L decreased browning intensity to 16%, 36%, 58%, 77% and 75% respectively. The study established that the best results for controlling browning were obtained when B. huillensis nodal segments were cultured on WPM medium supplemented with 5µM BAP while incorporated with 200 mg/litre of ascorbic acid (Figures 1(a) and (b)). Whereas, the study showed that
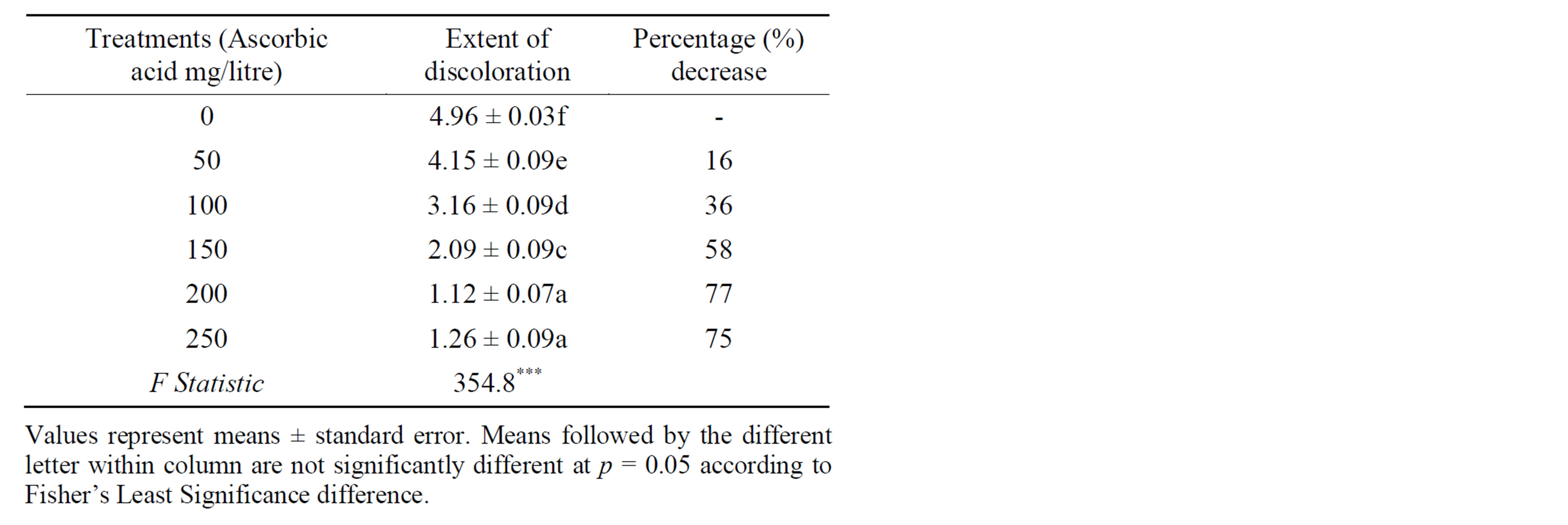
Table 1. Effects of ascorbic acid on lethal browning on nodal segments of B. huillensis.
 (a) (b)
(a) (b)
Figure 1. (a) & (b) = Nodal segments of B. huillensis (free from lethal brown) on WPM medium supplemented with 200 and 250 mg/litre ascorbic acid respectively.
the highest rate of coloration due to phenolic compounds was obtained in the culture medium without ascorbic acid (Figure 2(a)).
5. Discussion
Phenolic secretions and other exudates in plants tissue culture systems lessen explant initiation, growth, and development [18]. Therefore, preconditioning of explants with media supplements such as ascorbic acid is necessary to limit production of these substances [31].
In the current investigation, method for controlling lethal browning for nodal segments of B. huillensis was developed. The study established that the best results for controlling lethal browning were obtained when B. huillensis nodal segments were cultured on WPM medium supplemented with 5 µM 2, 4-D while incorporated with 200 mg/litre ascorbic acid. The result corroborates with the study conducted by [32] in woody plants and [33] in herbaceous plants (Musa spp). The ascorbic acid is able to scavenge oxygen radicals produced when the plant tissue is wounded and hence protecting the cells from oxidative injury [23]. Thus, as observed in this study, the ascorbic acid was useful and effective in managing the problem of browning caused by phenolic exudates and

Figure 2. (a) = Nodal segments of B. huillensis cultured on WPM medium free from ascorbic acid; (b) & (c) = Nodal segments of B. huillensis cultured on WPM medium supplemented with 50 and 100 mg/litre ascorbic acid respectively.
hence improving plant survival in vitro (Figures 1(a) & (b)); [20].
The results of the present study showed that almost all explants on the medium without ascorbic acid, and those with lower concentration levels of ascorbic acid browned extremely (Figures 2(a)-(c)). This may be attributed by the fact that exudation of phenolics is a natural mechanism in plants not only that but also many plants produce dark phenolic substances after wounding. Thus, accumulation of phenolic compounds in medium adversely affects the growth and survival of in vitro explants. [34,35] showed that accumulation of these compounds leads to browning and possibly death of the explants. Moreover, oxidized phenolic compounds may inhibit enzyme activity and result in the darkening of the culture medium and subsequent lethal browning of explants [36,37]. Thus, the study revealed that for successful control of oxidative lethal browning the concentration of antioxidant ascorbic acid is of great importance.
In conclusion, the problem of lethal browning has substantial constraint in in vitro propagation of B. huillensis. Nonetheless, we have manipulated the problem by incorporating the antioxidant (ascorbic acid) in woody plant medium. Besides that, significant difference between various concentration levels of ascorbic acid was noticed. The study established that the best result for controlling lethal browning was obtained when B. huillensis nodal segments were cultured on WPM medium supplemented with 5 µM BAP and incorporated with 200 - 250 mg/litre of ascorbic acid. Above all, the developed method is cheap and reproducible for combating oxidized lethal browning effect in B. huillensis nodal segments.
Acknowledgements
We thank the almighty whose blessings have enabled us to accomplish this work. Our sincere appreciation go to the Nelson Mandela African Institution of Science and Technology, the Commission for Science and Technology (COSTECH), Tanzania for financing the study, and Mikocheni Agriculture Research Institute (MARI) Dar es Salaam, for allowing us to use their laboratory facilities. We are also grateful to the Tanzania Forest Service (TFS) under the Ministry of Natural Resources and Tourism for permission to collect explant materials from BWFR.
NOTES