1. INTRODUCTION
Dendritic cells (DCs) are the most important antigen presenting cells (APCs) and, acting as sensors of microenvironment, capture pathogens and transmit the resulting information to lymphocytes. These activities can be carried out thanks to several types of highly specialized receptors such as C-Type Lectin Receptors (CLRs) and Toll like Receptors (TLRs) that are expressed on DC surface [1]. CLRs’ family acts in Ca2+-dependent manner [2], is able to bind and internalize carbohydrate structures and finally can induce the activation of T lymphocytes. Thanks to these properties CLRs are considered targets to enhance T cell-mediated protective immunity to prevent infectious diseases. Human MGL (Macrophage Galactose type C-type Lectin) is a C-type lectin expressed by immature myeloid Dendritic Cells (iDCs) and macrophages (Mfs), able to bind the N-Acetyl-Galactosamine residues (GalNAc or Tn). Any glycolipid, glycoprotein and pathogen carrying GalNAc is recognized and internalized by MGL expressing cells, thus including MGL in the list of Pathogen Recognition Receptors (PRRs) [3].
In the last years, it has been extensively demonstrated the involvement of this receptor in the uptake and processing of Tn-Tumor Associate Antigens (Tn-TAAs) [4,5]. In fact, MGL engagement acts as an adjuvant on DCs, promoting a phenotypic and functional maturation of the cells [4] and inducing Tn-TAAs cross-presentation [5]. The ability of MGL to stimulate the maturation of DCs and to induce a strong CD8+ T cell activation highlights the possibility to use this receptor as a target for anticancer vaccination.
Here we report the results obtained monitoring the expression of human MGL throughout a year on monocytes-derived DCs, in order to understand the optimal timing of MGL expression in DCs-based vaccination, with the purpose to use MGL as a vehicle for the delivery of Tn-TAAs.
2. MATERIALS AND METHODS
2.1. DC Generation
Peripheral Blood Mononuclear Cells (PBMCs) were isolated by Ficoll-Hypaque gradient (1077 g/mL; Pharmacia LKB, Uppsala, Sweden). Monocytes (CD14+) were purified by CD14 MicroBeads (Miltenyi Biotech, Paris, France) and cultured (5 × 105 cells/mL) in RPMI 1640 (Hyclone, UT, USA) supplemented with 2 mMLgluta-mine (Sigma Chemical Company, St. Louis, MO, USA), penicillin 100 U/mL (Sigma), streptomycin 100 μg/mL (Sigma) with 10% heat-inactivated Fetal Calf Serum (Hyclone). A total of 50 ng/mL rhGM-CSF (R & D System, Minneapolis, MN) and 1000 U/mL rhIL-4 (R & D System) were added at day 0 and 2. iDCs were collected at day 5.
2.2. DC Phenotype
Cell phenotype staining was performed using the following antibodies: MoAbs anti-hMGL (ASGPR/MGL, 125A10.03 clone, Dendritics, Lyon, France) followed by FITC-conjugated goat anti-mouse IgG (H + L) (Jackson ImmunoResearch Laboratories, West Grove, PA, USA). MoAbs directly conjugated with FITC or phycoerythrin (PE) were also employed: IgG-FITC and -PE as isotype controls, anti-HLAII-DR-FITC, anti-CD86-FITC, antiCD83-PE, anti-CD40-PE, anti-CD14-PE all from Becton Dickinson (San Diego, CA, USA) and anti-CCR7-FITC from R&D. DCs were incubated with conjugated-MoAb for 30 min at room temperature (RT) as indicated by the manufacturer’s instruction. After washing, at least 1 × 104 events were evaluated using a FACSCanto flow cytometer running FACSDiva acquisition and data analysis software (Becton Dickinson).
3. RESULTS
We monitored the MGL expression on iDCs derived from 144 healthy donors throughout a year (12 donors/ month). Monocytes were differentiated for 5 days in iDCs (Figure 1(a)) and the presence of the receptor was analysed by cytofluorimetry. The results obtained showed that MGL expression on iDCs surprisingly had a seasonal modulation: the expression was higher in winter rather than in summer, despite that yield and quality of DCs were homogeneous. Figure 1(b) shows the expression of MGL as fold increase, obtained by the ratio between the Mean Fluorescence Intensity (MFI) of DCs stained with an anti-MGL antibody and the MFI obtained by the addition of the isotype control. We considered MGL+ donors all those samples that showed at least a two fold increase. As described in Figure 1(c), MGL expression among donors displayed a negative linear trend going from the winter (100% in January; 50% in March) to the spring dropping to 33% during the summer period. The percentage of MGL+ donors began to increase in September (58%) reaching the 83% at the end of the year (December).
4. DISCUSSION
In this report we showed for the first time a circa-annual rhythm of human MGL receptor on iDCs. Its expression was higher in winter rather than in summer, as well as several C-Type Lectins expressed by other organisms such as plants [6].
 (a)
(a)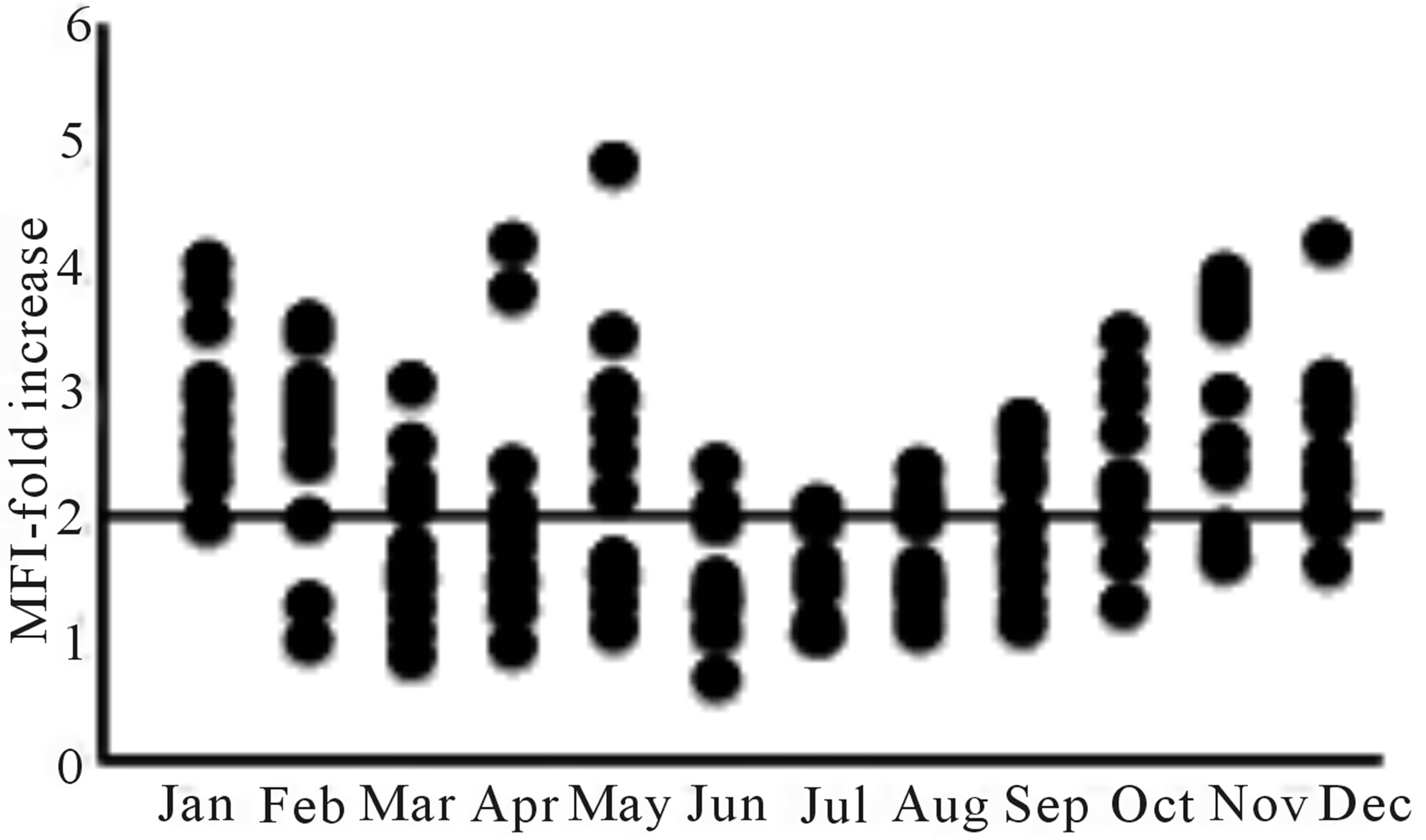 (b)
(b) (c)
(c)
Figure 1. Expression and modulation of the MGL receptor on DCs: (a) iDC phenotype at day 5 of differentiation analyzed by cytofluorimetry. The empty histograms represent the isotype control. The panel shows a representative donor; (b) MGL expression evaluated by cytofluorimetry adding an anti-MGL antibody (125A10.03 clone) or the IgG1 isotype control to the iDCs followed by Fluorescein Isothiocyanate (FITC)-conjugated goat anti-mouse IgG (H + L). For each donor, MGL expression was reported as fold increase of the MFI obtained with the anti-MGL antibody / MFI of the isotype control. All donors with a ratio ³ 2 were considered MGL+; (c) Percentage of MGL+ donors throughout a year.
MGL represented an exclusive marker of myeloid iDCs and it has been demonstrated in vitro that its expression was increased by dexamethasone (Dex) treatment [7]. Up to now little is known on the factors that can modulate the circa-annual rhythm of the human lectins [6,8] on DCs, but extensively studies on the impact of glucocorticoids (GCs) like Dex on DCs were conducted. Rea D. et al. showed that Dex affected the viability of DCs, selectively down-regulated the expression of costimulatory molecules reducing their immunostimulatory properties [9]. Moreover in vivo studies showed that a single injection of Dex impaired DC antigen presentation [10]. These results indicated that GCs regulated the DC maturation and immune functions in vitro and in vivo, demonstrating that GCs played a role in the prevention of spontaneous activation of DCs, decreasing their T-cell stimulatory potential and influencing the expression of molecular markers, such as MGL. Furthermore GCs in human seemed to have per se a seasonal modulation [11], that was similar to that observed for MGL. These variations were probably due to the stress-related factors that were differently released during the year. These evidences prompted us to believe that GCs could be one of the possible factors that contribute to MGL circa-annual rhythm.
Interestingly, similar seasonal modulation has been demonstrated for other PRRs, such as TLR (Toll-like Receptor)-2 and -4. Their expression on PBMCs was shown to be seasonally modulated by blood circulating Vitamin D3 resulting in the secretion of distinct cytokine patterns that contribute to alter the innate immune response [12].
The modulation of MGL on DCs as well as of other PRRs might be due to seasonal changes of immune system probably characterized by several molecular mechanisms contributing to the response of immune system to the seasonal stress stimuli, even if further studies need to be performed in order to identify all the processes implicated in this phenomenon. Since MGL is a PRR with the capacity to internalize its ligands and has adjuvant effects on DCs, this modulation can have important implications in the planning of vaccines against pathogens as well as in the identification of the optimal timing for DCs-based vaccination in cancer patients.
5. ACKNOWLEDGEMENTS
This work was supported by Ministero per l’Istruzione e la Ricerca Scientifica (AR: PRIN 2009NREAT2_004, MN: PRIN 2009XMZPKW), AIRC IG2009, “Sapienza”, Università di Roma and Ministero della Salute (AR, MN, CN: C26AI2L3RA).