Oxidation of Amino Acids by Chlorpromazine Cation Radical and Co-Catalysis by Chlorpromazine ()
1. Introduction
Chlorpromazine (CPZ) is a neuroleptic drug of the phenothiazine family and is widely used in the treatment of schizophrenia [1]. However, the long-term use may cause severe side effects [2], which in many cases have been related to oxidative damage. Regarding hepatotoxicity, which is one of the major concerns during the clinical use of CPZ, this neuroleptic drug is able to inhibit intracellular carboxylesterases, such as alpha-naphthyl acetate esterase, naphthol chloroacetate esterase, and alphanaphthyl butyrate esterase in hepatocytes [3]. Melatonin, a largely used antioxidant, when administered with CPZ, significantly attenuated the oxidative stress parameters as evidenced by lowering malondialdehyde levels in tissue homogenate [4]. In the brain, the level of the lipid peroxidation product, hydroxyalkanals, is increased when the rats are treated of CPZ [5].
The above chemical features of CPZ might be related to the pro-oxidant effect of chlorpromazine cation radical (CPZ•+), which can be easily generated by catalytic action of peroxidases, including the neutrophil myeloperoxidase (MPO) and by methemoglobin [6,7]. For instance, the in vitro involvement of CPZ•+ during the oxidation of biomolecules has been well-established for catecholamines, ascorbate, GSH, NADH and melatonin [8-10]. Additionally, enzymes are inactivated by CPZ•+. This is the case for cholinesterase in rat serum [11], and for trypanosoma cruzi dihydrolipoamide dehydrogenase, through its interaction with phenothiazine radicals generated by MPO or horseradish peroxidase (HRP)/H2O2 systems [12].
For the reasons stated above, here we aimed to study the reactivity of CPZ•+ with amino acids. The experimental approaches were the direct reaction between CPZ•+ and amino acids and the co-catalysis effect of CPZ when the amino acids were submitted to oxidation by HRP/ H2O2.
2. Materials and Methods
2.1. Chemicals
Tryptophan, tyrosine, cysteine, methionine, glycine, phenylalanine, aspartic acid, lysine, uric acid, NaNO2, ascorbic acid, 5,5’-dithiobis(2-nitrobenzoic) acid (DTNB), horseradish peroxidase (HRP) (EC 1.11.1.7) and chlorpromazine were purchased from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA). Hydrogen peroxide (H2O2) was prepared by diluting a 30% stock solution and calculating its concentration (ε240 nm = 43.6 M–1×cm–1). All the reagents used for buffers and mobile phase were of analytical grade.
2.2. Oxidation of Amino Acids by Pre-Formed CPZ•+
Solutions of CPZ•+ were prepared as follows: CPZ (1 mM) was dissolved in pH 3.8 glycine buffer (20 mM). Then, 0.1 μM HRP and 100 μM H2O2 were added and the reaction was allowed to proceed to completion (1 min). The initial concentration of CPZ•+ was calculated using its extinction coefficient ε525 nm = 12.100 M–1×cm–1 [13]. Considering the absorbance obtained of 1.4, the initial concentration of CPZ•+ was 115 μM. The amino acids were added 20 s after the formation of CPZ•+. The experiments were monitored using a Hewlett Packard 8452 diode array spectrophotometer.
For the HPLC studies, the reaction mixtures were incubated for 10 min at 25˚C and injected into the HPLC system. Determinations of tryptophan and tyrosine consumptions were carry out using HPLC combined with diode array detection. The separation was performed on a 15 × 6.0 mm i.d., 5 μm, 100 Å, C18 Shin-pack column (Shimadzu, Kyoto, Japan). The HPLC system (Jasco, Tokyo, Japan) consisted of a PU-2089s quaternary gradient pump, a CO-2060 column oven set at 25˚C and a MD-2015 diode array detector. The reaction mixtures were separated were separated by using a gradient of water (solvent A) and methanol (solvent B), programmed as follows: 0 - 10 min from 95 to 70% A, 10 - 15 min from 70% - 95% A. The total flow rate of the mobile phase was 1.0 mL/min and the injection volume was 20 μL. The diode array detector wavelength of 260 nm was used to monitor tryptophan consumption and 230 to monitor tyrosine consumption.
2.3. Oxidation of Tryptophan and Tyrosine: Co-Catalysis by CPZ
In these experiments CPZ•+ was not pre-formed as above. The reaction mixtures were constituted by tryptophan 50 μM or tyrosine 50 μM, HRP 0.1 μM, CPZ 10 μM, H2O2 100 μM in 10 mM PBS pH 7.4 at 25˚C in the presence or absence of the alternative substrates. The reactions were triggered by adding H2O2 and incubated by 10 min. After oxidation, the remaining concentration of tryptophan and tyrosine were analyzed by HPLC as above.
2.4. Oxidation of Tryptophan: Comparison between Co-Catalysis by CPZ and Oxidation Using High Concentration of H2O2. Consumption of Tryptophan and Formation of Products
Using CZP as a co-catalyst the reaction mixtures were constituted by 1 mM tryptophan, 2 mM H2O2, 100 μM CPZ and 1 μM HRP in PBS pH 7.4 at 25˚C and incubated for 10 min. Using high concentration of H2O2, the oxidation was performed as previously described, as follows: reaction mixtures were 1 mM tryptophan dissolved in 0.1 M borate buffer pH 6.3. H2O2 was added hourly (final concentration of 1.0 M) for 6 h at 37˚C in dark as previously reported [14]. In both methods the separation of oxidation products was carry out isocratically on a 15 × 6.0 mm i.d., 5 μm, 100 Å, C18 Shin-pack column, using a mixture of solvents composed of 0.1% acetic acid (94%) and methanol (6%). The total flow rate of the mobile phase was 1.0 ml/min and the injection volume was 20 μl.
2.5. Oxidation of Cysteine and Co-Catalysis by CPZ
Cysteine (1 mM) was incubated with 0.1 μM HRP, 100 μM H2O2 in 10 mM PBS pH 7.4 at 25˚C in the presence or absence 25 μM CPZ. The reactions were triggered by adding H2O2 and stopped at regular intervals by adding 20 μg/ml catalase. The remaining concentration of cysteine was analyzed by the reduction of DTNB as follows: 450 μl of the reaction mixture was added to 450 μl of 300 mM Na2HPO4 and 100 μl of DTNB solution (20 mg DTNB in 100 ml 1.0% sodium citrate). The absorbance was read at 412 nm using PBS as blank [15]. A standard curve was generated for calculation of cysteine concentration.
3. Results
The stability of CPZ•+ is strongly dependent of the identity, concentration and pH of the buffer. Here, we used 20 mM pH 3.8 glycine buffer, where CPZ•+ attained its higher lifetime [16]. CPZ•+ was prepared through oxidation of CPZ catalyzed by HRP, where the cation radical was stable for at least 20 min. The concentration of CPZ•+, calculated based in its excitation coefficient at 525 nm, was about 100 μM [13].
To study the reactivity between the amino acids and CPZ•+, these compounds were added in the pre-formed CPZ•+. Figure 1 shows that only tryptophan, tyrosine and cysteine, provoked a fast decay in the concentration of CPZ•+. Lysine, aspartic acid, methionine and phenylalanine were unreactive. The kinetic experiment also revealed a higher reactivity between CPZ•+ and tryptophan

Figure 1. Effect of amino acids and nitrite on CPZ•+. CPZ•+ was pre-formed as described in materials and methods. (a) UV-Vis absorption band of CPZ•+ and its bleaching by tryptophan. The scans were taken every 20 s. (b) Decay of CPZ•+ provoked by amino acids or nitrite (50 μM). The substrates were added 40 s after the formation of CPZ•+.
when compared to tyrosine.
The spontaneous bleaching of CPZ•+ is caused by its reaction with the buffer components, including water and other nucleophiles, which lead to the parent CPZ and CPZ-sulfoxide [16]. Here, searching for biomolecules that could react with CPZ•+, we found that nitrite anion (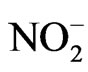 ), which is present in high concentration in the blood plasma, increased significantly the decay of CPZ•+ (Figure 1). However,
), which is present in high concentration in the blood plasma, increased significantly the decay of CPZ•+ (Figure 1). However,  was not consumed during the reaction. The analysis of
was not consumed during the reaction. The analysis of  was performed by measuring its concentration before and after the bleaching of CPZ•+ (Griess method, not shown).
was performed by measuring its concentration before and after the bleaching of CPZ•+ (Griess method, not shown).
Considering that some compounds provoke the bleaching of CPZ•+ but are not oxidized, the concentration of amino acids were measured after the reactions. The results in Tables 1-2 show that CPZ•+ was able to oxidize these amino acids. The results also show that others substrates, which are normally present in the blood plasma, were able to compete and partially inhibit the oxidation of tyrosine and tryptophan. Additionally, the results in Tables 1-2 show the higher reactivity of CPZ•+ with tryptophan when compared to tyrosine and confirm the kinetic experiments. How depicted in the tables, tryptophan inhibited the oxidation of tyrosine in 59%, while tyrosine inhibited the oxidation of tryptophan in only 18%.
Besides the oxidizing capacity of CPZ•+, it was also found that CPZ is able to act as a co-catalyst during the HRP-catalyzed oxidation of amino acids by H2O2. In these experiments, CPZ•+ was not pre-formed, but generated during the reaction. The data in Table 3 show that tryptophan was completely unreactive under this experimental condition, but was almost completed oxidized in the presence of CPZ, which was used in a concentration 5 times lower than tryptophan. Similar results were obtained to tyrosine (Table 4). The inhibitory effect of alternatives substrates upon oxidation of tryptophan and tyrosine was also studied using CPZ as a co-catalyst. In this case the inhibitory effect was significantly lower compared to the experiments with pre-formed CPZ•+.

Table 1. Oxidation of tryptophan by pre-formed CPZ•+ and effect of alternative substrates.

Table 2. Oxidation of tyrosine by pre-formed CPZ•+ and effect of alternative substrates.

Table 3. Oxidation of tryptophan by HRP/H2O2 and cocatalysis of CPZ: effect of alternative substrates.

Table 4. Oxidation of tyrosine by HRP/H2O2 and co-catalysis of CPZ: effect of alternative substrates.
The co-catalytic effect of CPZ was studied by UV-Vis measurements and demonstrated the concentration dependent effect of CPZ on HRP-catalyzed oxidation of tryptophan. Similar results were observed by monitoring the decay of native tryptophan fluorescence (Figure 2).
The efficiency of tryptophan oxidation using CPZ as a co-catalyst was compared with a recently reported method, which is based in the use of high concentration of H2O2 (1 M) in borate pH 6.3 buffer [14]. Using the method developed here and 1 mM of tryptophan, the consumption of tryptophan was 0.71 ± 0.05 mM and the reaction was completed in 15 min. By using the reported method, after an incubating for 6 h, the consumption was 0.12 ± 0.05 mM. These are mean and SD of three experiments. Figure 3 shows the chromatograms of the reaction mixtures using both methods. The products (peaks) were also similar in both methods.
CPZ was also an effective co-catalyst to cysteine. In this case, after the incubation with HRP and H2O2, the remaining concentration of cysteine was measured by the reduction of DTNB (Figure 4).
4. Discussion
The inhibitory effect of CPZ•+ upon enzyme activity [11,12] is probably related to protein oxidative damage. Here, our results reinforce this proposal since CPZ•+ oxidized easily the amino acids tryptophan, tyrosine and cysteine. As well established in the scientific literature, the oxidation of these amino acids has been implicated in the loss of functions of protein and has been detected in physiologically important proteins [17-19]. Here, special attention was given to tryptophan, since this amino acid was more reactive than tyrosine with CPZ•+. The higher reactivity between CPZ•+ and tryptophan could be related
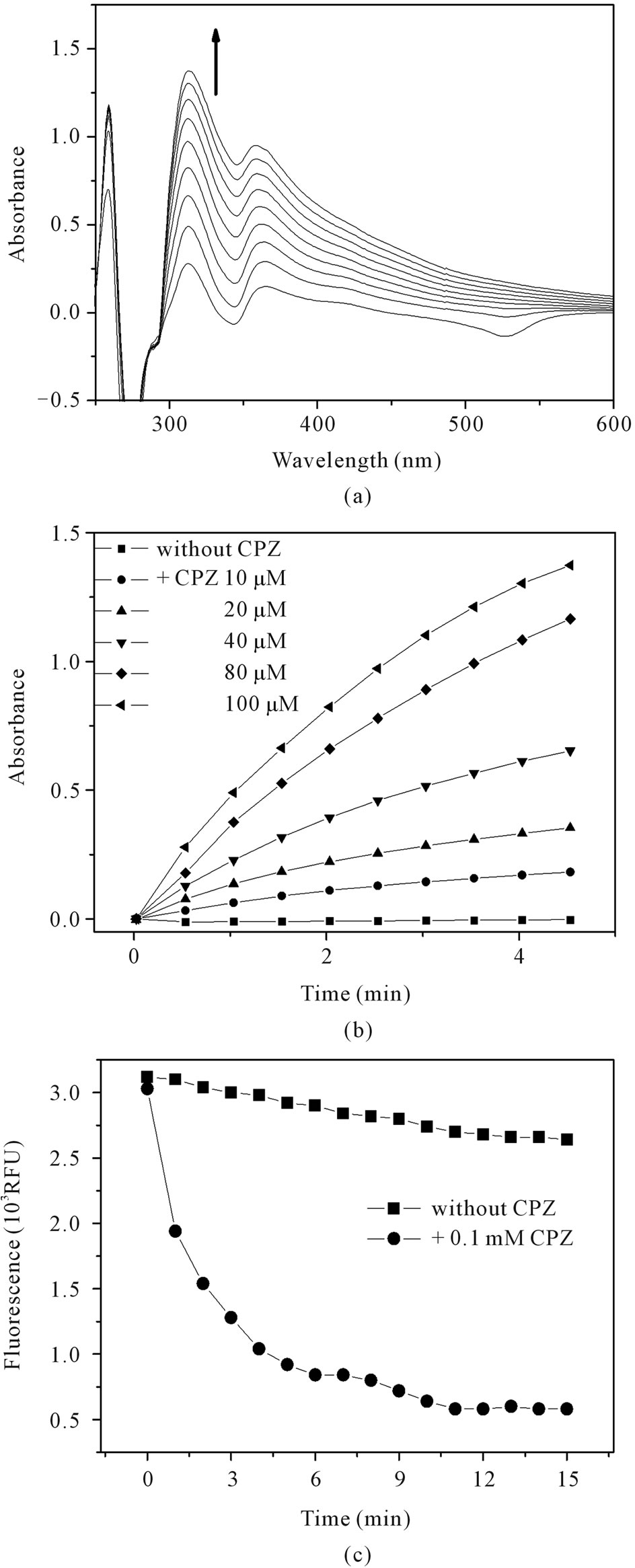
Figure 2. Oxidation of tryptophan: Co-catalysis by CPZ. (a) UV-Vis difference spectra obtained during oxidation of tryptophan. The scans were taken at 30 s intervals. (b) Concentration-dependent effect of CPZ on tryptophan oxidation. (c) The oxidation of tryptophan monitored by its native fluorescence (εex = 290, εem = 360). The reaction mixture contained 1 mM tryptophan, 2 mM H2O2, 1 μM HRP in PBS pH 7.4 at 25˚C.
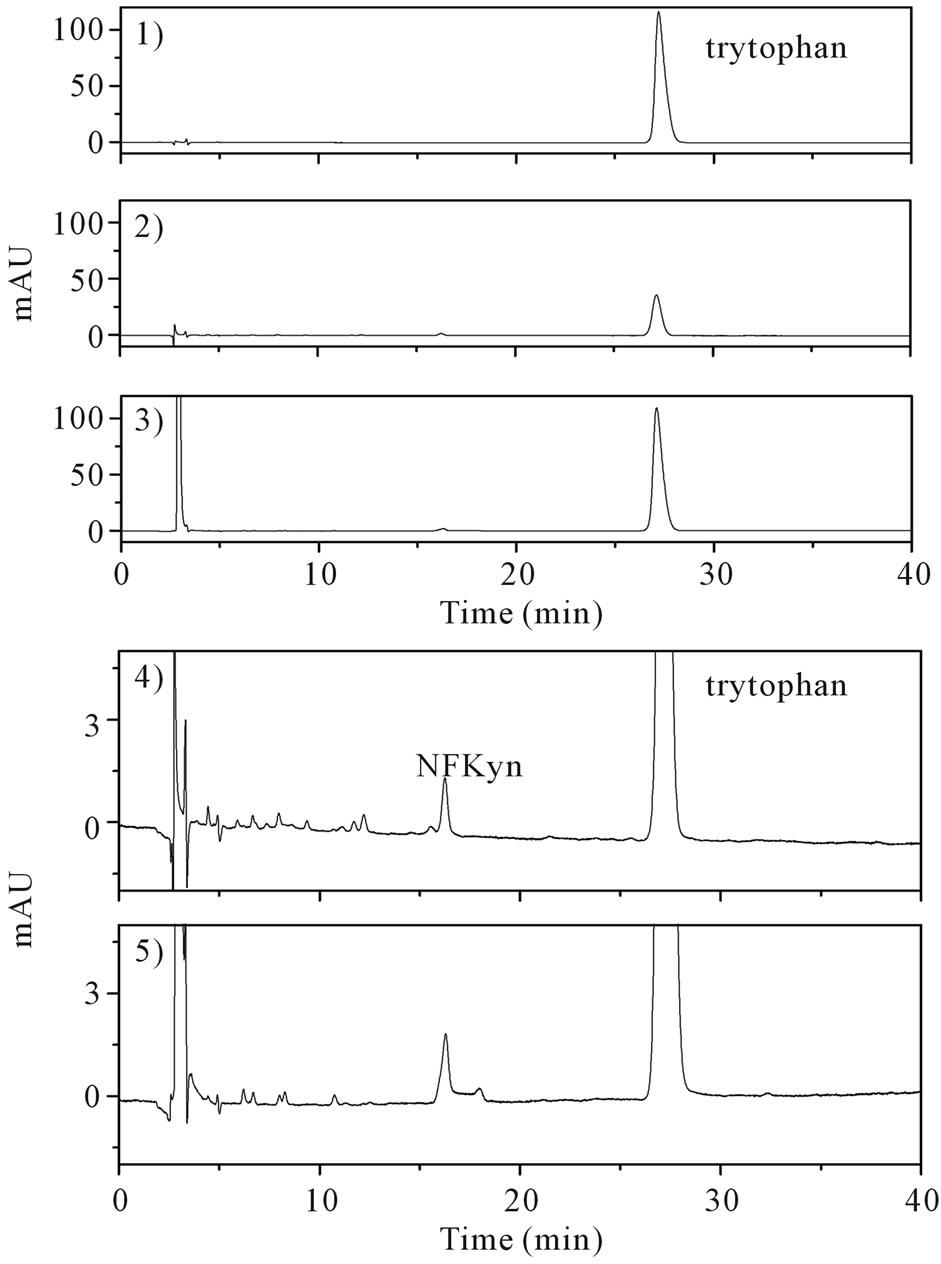
Figure 3. Efficiency of oxidation of the tryptophan: comparison between co-catalysis by CPZ and oxidation by high concentration of H2O2. 1) Standard of tryptophan 1 mM; 2) Remaining tryptophan, 10 minutes after the beginning of the reaction using CPZ. The reaction mixture contained 1 mM tryptophan, 2 mM H2O2, 1 μM HRP, 100 μM CPZ and in PBS pH 7.4 at 25˚C; 3) Remaining tryptophan, 6 hour after the beginning of the reaction using 1 M H2O2. The reaction mixture contained 1 mM tryptophan and 1 M H2O2; 4) Products using CPZ (expanded chromatogram 2). 5) Products using 1 M H2O2 (expanded chromatogram 3). N-formylkynurenine (NFkyn).
to favorable interaction between this amino acid and CPZ as have been verified by suppression of fluorescence of serum albumin by CPZ [20]. In the same direction, the inactivation of cholinesterase induced by CPZ•+ was correlated with depletion of tryptophan residues [13].
Considering the formation of CPZ•+ and its putative deleterious consequences in the physiological medium, it should be also consider the presence of other oxidizable metabolites that could interact and neutralize this oxidant. For this reason, we compared the relative efficacy of oxidation of tryptophan and tyrosine when compound as nitrite, ascorbate, cysteine and uric acid, which are natural components from blood plasma, were added in the reaction mixture. The significant inhibition in the oxidation of tryptophan and tyrosine obtained when these alternative metabolites were added in the reaction mixture is an indication that CPZ•+ can be reduced by many other

Figure 4. Oxidation of cysteine: Co-catalysis by CPZ. The reaction mixture (control) contained 1 mM cysteine, 100 μM H2O2, 0.1 μM HRP in PBS pH 7.4 at 25˚C. When present 25 μM CPZ. The remaining concentration of cysteine was measured with DTNB. The results are mean and SD of triplicates.
oxidizable and non-oxidizable endogenous compounds. The last case is applicable to nitrite, since this metabolite is not oxidized by CPZ•+, but increased its rate of spontaneous decomposition and, as a consequence, inhibited the oxidation of tyrosine and tryptophan. However, the efficacy of the alternative endogenous substrates used here as inhibitor of tryptophan and tyrosine oxidations was significantly lower when CPZ acted as a co-catalyst. For instance, the effect of nitrite could be considered, as it caused strong effect on CPZ•+ stability. In the experiments with pre-formed CPZ•+ the inhibition of oxidation was 73% and 100% for tryptophan and tyrosine, respectively. In the experiments using CPZ as a co-catalyst, the inhibitions were only 14% and 4%, respectively. The main difference between these experiments is that using CPZ as a co-catalyst, CPZ•+ acts as a transient intermediate, that is not accumulated during the reaction course. A mechanism proposal, for the lower inhibitory effect of nitrite in this case, would be an interaction between CPZ•+ and tryptophan or tyrosine close the active site of HRP. In other words, CPZ•+ would not diffuse into the bulk solution. Finally, since the action of CPZ as a co-catalyst is a condition closer that could be found in the physiological medium, we propose, that the oxidation of amino acids can really take place and be involved in proteins damage when patients are chronically submitted to treatment with this neuroleptic drug. The scheme depicted in Figure 5 presents a proposal for the involvement of CPZ during the oxidation of amino acids.
Considering yet the effect of CPZ as a co-catalyst during tryptophan oxidation, it must be considered that this amino acid is particularly unreactive with HRP/H2O2.

Figure 5. Proposal for the involvement of CPZ as a cocatalyst during the oxidation of amino acids.
Indeed, the oxidation of indole derivatives by H2O2, and catalyzed by peroxidases as HRP or MPO, is dependent of the reduction potentials of these compounds. For tryptophan (E˚ = 1.015 V), the bimolecular rate constant for its reaction with the active form compound-II (HRP-II) is 6.9 ± 1.3. For melatonin (E˚ = 0.95 V), the rate constant increase to (9.6 ± 0.3) × 102. For additional comparison, tyrosine a phenolic compound (E˚ = 0.93), the rate constant reached (1.6 ± 0.6) × 104 [21]. Here, this poor reactivity of tryptophan was confirmed when 1 mM was submitted to oxidation with 2 mM H2O2 and 1 μM HRP. In this case, our results demonstrated that no more than 18 μM was oxidized in such condition. However, in the presence of CPZ as a co-catalyst the uptake reached 710 μM.
The comparison between the efficiency of tryptophan oxidation using our method (tryptophan 1 mM, H2O2 1 mM, HRP 1 μM and 0.1 mM CPZ), where 710 μM of tryptophan was consumed in 10 min, with the described method, that use high concentration of H2O2 (1 mM tryptophan and 1 M H2O2), where the consumption was 120 μM after 6 h [14], demonstrated the efficacy of CPZ as a co-catalyst. It was not our interest the identification of tryptophan’s oxidation products, however, a comparison of the chromatogram peaks profile, with the reported method above, shows that similar products were obtained using CPZ. Among these, it was found a peak that has retention time and UV-Vis spectrum (not shown) that matched the product of oxidative cleavage of the indole ring, N-formylkynurenine.
Besides tryptophan and tyrosine, and how could be expected for a sulfhydryl compound, cysteine was also easily oxidized by CPZ•+ and by CPZ as a co-catalyst. This is clear evidence and suggests that proteins could be cross-linked by interacting with this neuroleptic drug.
In conclusion, the high reactivity of oxidizable amino acids with CPZ confirms the potential deleterious effect of this drug regarding protein damages. The particular efficacy of tryptophan oxidation using CPZ, when compared to oxidation mediated by peroxidase alone, could be a useful method to produce oxidized tryptophan residues in protein. Indeed, there is a current interest in the development of analytical procedure that could detect oxidized tryptophan in proteins [22,23]. Hence, using the method developed here, the production of protein with oxidized tryptophan could easily achieve.
6. Acknowledgements
This work was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (Fapesp) and from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).