Received 18 November 2015; accepted 12 January 2016; published 15 January 2016

1. Introduction
The selective depletion of CD4+T cells, the cells commonly known as helper T cells or T4 cells by the human immunodeficiency virus (HIV) is the cause of infection in human and has been the subject of most intense studies that encompass diverse field of scientific studies. HIV infect CD4+T cells which play a central role in immune regulation and their depletion can have widespread deleterious effects on the functioning of the immune system as a whole, this is cause for alarm and a key reason for HIV devastating effect [1] . Indeed, the decline in the number of CD4+T cells in peripheral blood is used in a clinical setting as an indicator of the disease stage [2] . The puzzle today is that while much progress has been achieved by medical and biological researchers in understanding aspect of the virus-host interaction, the mechanism by which HIV causes AIDS still remains unexplained.
The dynamics of immune response to any virus involves different components and is generated by a complex web of interactions among different types of white blood cells (monocytes, T and B cells). Although they can be variations from one patient to another, the time scale to develop a specific immune response may vary from days to weeks. In the case of HIV, the entire course of infection involves two different time scales [3] . The primary infection which shares many similarities with acute infections, exhibits the same characteristics as any other viral infection: a dramatic increase of the virus population during the first 3 - 6 weeks, followed by a sharp decline, due to the action of the immune system. Instead of being completely eliminated after the primary infection, as many other viruses, however, a low HIV concentration is detected for a long asymptomatic time: the clinical latency period. During this period, the immune response keeps the viral load to a constant level referred to as the set point viral load [4] . This period may vary from one to ten (or more) years [5] . Besides the low virus burden detected during this period, a gradual deterioration of the immune system is manifested by the reduction of CD4+T cell populations in the peripheral blood. The third phase of the disease is achieved when the concentration of the T cells is lower than a critical value (~30%), leading to the development of AIDS.
Differential equations represent good approximations to situations where there are large number of events happening on average and when the time scale over which the average is being examined is much longer than the interval between events. At the initial time when the research into HIV/AIDS started, it was not understood that in HIV-infected individuals the virus was being produced at a prolific rate even before the onset of symptoms and full-blown AIDS. Mathematical models have been proven valuable in understanding the dynamics of HIV infection. Perelson and Nelson [5] developed a simple model for the primary infection with HIV. This model has been important in the field of mathematical modeling of HIV infection, and many other models have been proposed, which take this model as their inspiration. Perelson et al. [6] extended the model in 1993 and discussed some of the model's behavior. They defined the model by considering four categories: uninfected CD4+T cells, latently infected CD4+T cells, productively infected CD4+T cells and virus population. Wodarz and Nowak [7] used their model to measure crucial parameters which led to a new understanding of the disease process. Furthermore, in their work they showed how mathematical models can be used to understand correlates of long-term immunological control of HIV, and to design therapy regimes that convert a progressing patient into a state of long-term non-progression. Shi, V. et al. [8] formulated a novel cellular automated (CA) model for HIV dynamics and drug treatment. The model is built upon realistic biological processes, including the virus replication cycle and mechanism of drug therapy. Viral load, its effect on infection rate, and the role of latently infected cells in sustaining HIV infection are among the aspects that are explored and incorporated in the model. The results of the simulation show the three phases of HIV dynamics. In the rest of what follows, we introduce our mathematical model of HIV without treatment in Section 2. In Section 3, we introduce a linearization technique. Section 4 deals with the numerical simulation and analysis. The last section is the conclusion.
Statement of Problem
Nonlinearity can be classified as inherent (natural) and intentional (artificial). The case of interaction of HIV with the immune system is one that has undesirable effects; control and eradication have been of increasing concern to clinicians and mathematical modellers. The behaviour of this nonlinear system, however, is much more complex as HIV evolves and behaves differently from other viruses. Can there then be any equilibrium point?
2. HIV Model without Treatment
We consider the model developed by Kirschner, D and Webb [4] , in which Pritikin [9] obtained a numerical solution showing the course of HIV. Ogban and Lebedev [10] carried out numerical stability of [9] . The objective here is to seek physical stability (analytic solution) to the system. Linearizing the system, we compare our analytic solution with the numerical solution obtained in [10] . We introduce a system of ordinary differential equations modeling the immune dynamics of an HIV infected immune system in the absence of treatment. The variables are defined as follows:
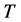 ―Concentration of uninfected CD4+T cells;
―Concentration of uninfected CD4+T cells;
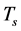 ―The concentration of CD4+T cells infected with HIV;
―The concentration of CD4+T cells infected with HIV;
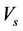 ―Concentration of HIV virus.
―Concentration of HIV virus.
We note that the model without treatment describes the processes in the blood, the replication of the virus and mortality of cells occurring in the lymphatic system, and as a result, the model describes the dynamics observed in the blood variables rather than operating characteristics of infection.
The derivatives with respect to time of these variables satisfy the system of differential equations:
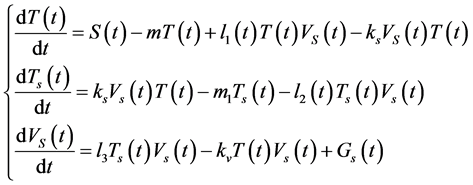 (1)
(1)
The system of equation has the initial conditions:
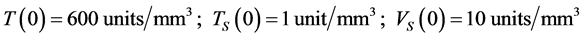
The expressions on the right sides of Equations (1)-(3) indicate the following: In Equation (1), 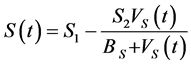 is a function, which represents the source of, uninfected CD4+T-cells from the thymus
is a function, which represents the source of, uninfected CD4+T-cells from the thymus
and other compartments. Here 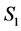 and
and 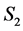 are constants,
are constants, 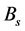 is a saturation constant (saturation ratios introduced into the model to adjust the parameters of growth under great changes in populations during the course
is a saturation constant (saturation ratios introduced into the model to adjust the parameters of growth under great changes in populations during the course
of infection and treatment). m-is the mortality rate of uninfected CD4+T-cells (birth rate =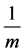 );
); 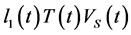 where
where 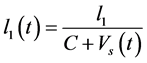 and C describes the proliferation rate of CD4+T-cells in the plasma
and C describes the proliferation rate of CD4+T-cells in the plasma
eliciting an immune response, due to the effect of stimulating the immune system antigen; this explains the increased turnover member of CD4+T-cells;
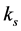 ―is the infection rate of CD4+T-cells by the virus.
―is the infection rate of CD4+T-cells by the virus.
In Equation (2):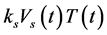 ―the growth rate of infected CD4+T-cells as the virus infects T-cells;
―the growth rate of infected CD4+T-cells as the virus infects T-cells;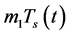 ― a loss due to mortality of the infected cells;
― a loss due to mortality of the infected cells;
![]() where
where ![]() is a saturation coefficient describes the death of infected cells owing to the presence of the virus.
is a saturation coefficient describes the death of infected cells owing to the presence of the virus.
In Equation (3): The virus population increases due to the term ![]() where
where ![]() this term describes the increase in the population of virus in the blood. The dependence of this term on
this term describes the increase in the population of virus in the blood. The dependence of this term on ![]()
takes into account the reduction in the proliferation of the virus in the plasma when the concentration of infected CD4+T cells in the plasma decreases. Since most virus enters into the plasma from the external source of lymph, the plasma viral population during the final stage of the infection grows rapidly; ![]() -describes the
-describes the
destruction of the virus by the immune system; ![]() (where B saturation constant) takes into
(where B saturation constant) takes into
account the entry of the virus from the lymphoid system. This term is a major contributor to the population of virus in the blood. The parameter values are as given in Table 1, see [10] .
![]()
Table 1. List of constants and parameters.
3. Linearization Technique
The similarity of a nonlinear system to a linear system in the local region of each singular point can be formalized by linearizing the nonlinear system as we now discuss.
If the singular point of interest is not the origin, by defining the difference between the original state and the singular point as a new set of variables, one can shift the singular point to the origin. Therefore, without loss of generality, we may simply consider Equation (1) with a singular point (0, 0, and 0) [6] . Thus (1a-c) can be rewritten as a system
![]() (2)
(2)
We can write (2a-c) as
![]() (3)
(3)
where![]() ,
, ![]() ,
,![]() .
.
Thus, if we write
![]() (4)
(4)
where (3) is equal to (4) if g(t) is neglected. By [11] , the nonlinear system (3) is a perturbation of the associated linear system (4). Now for system (2) the Jacobian matrix is given as
![]() (5)
(5)
We find that this system has one equilibrium point![]() . The Jacobian at the fixed point
. The Jacobian at the fixed point ![]() is
is
![]() (6)
(6)
The characteristic equation of (6) is given by
![]() (7)
(7)
![]() (8)
(8)
Since the system is three-dimensional, we use the Routh-Hurwitz criterion to establish the negativity of the real parts of the roots of the characteristic equation and therefore stability of the equilibrium under consideration. For a three-dimensional system see [12] [13] , with a characteristic equation of the form:
![]()
The Routh-Hurwitz criteria state that all roots of the characteristic equation have negative parts (local stability) if and only if ![]()
Analysis of the characteristic equation evaluated at the fixed point, shows that it is unstable. It is however stable if![]() .
.
4. Numerical Simulation and Analysis
Using MathCAD, we employed Runge Kuta method of order 4 to obtain our simulations. It has been reported that during acute infection the individual become antigenic and viremic. High levels of infectious virus can be detected in the peripheral blood [14] . With the initial conditions as given above, we investigate the influence of the viral reproduction rate on the three variables. Our calculations with the same initial conditions are shown to be consistent with results from the authors in [4] [9] . This fact validates our computational work with the models which are without treatment in [4] [9] . In [4] [9] , they showed that remission depended on a certain threshold of the virus level.
Figures 1-3 are the numerical solutions to Model 1. The parameter values used to generate these figures can be found in Table 1 [10] . The simulations of the dynamics are with T(0) = 600/mm3, Ts(0) = 0, Vs(0) = 10. The viral reproduction rates are: ![]() and
and ![]() Noting that the model is without treatment, if the parameter value is kept very low then it takes longer time for the level of T-cells to get lower than 200/mm3. This means that a low value of the parameter does not push the system into the progression to AIDS. In other words, the steep crash in day 3000 takes longer to achieve. This suggests that with low levels reproduction of virus the infection may go extinct rather than take off, see [4] [5] [9] [10] . Complete inhibition of viral replication appears
Noting that the model is without treatment, if the parameter value is kept very low then it takes longer time for the level of T-cells to get lower than 200/mm3. This means that a low value of the parameter does not push the system into the progression to AIDS. In other words, the steep crash in day 3000 takes longer to achieve. This suggests that with low levels reproduction of virus the infection may go extinct rather than take off, see [4] [5] [9] [10] . Complete inhibition of viral replication appears
![]()
Figure 1. Graphical simulations of uninfected CD4+T cells against time.
![]()
Figure 2. Graphical simulations of infected T cells against time.
![]()
Figure 3. Graphical simulations of virus cells against time.
impossible and may be unnecessary. In Figures 1-3, the curves ![]() represent the values
represent the values ![]() and
and ![]() respectively.
respectively.
As shown in [4] , the curves of Figures 1-3 are consistent with the results of clinical trials of HIV [15] - [18] . After a period of acute infection during the first few weeks after seroconversion, the number of CD4+T cells gradually decline from approximately from ![]() to zero over a period of time equal to approximately 10 years (normal number of CD4+T cells varies in the range of
to zero over a period of time equal to approximately 10 years (normal number of CD4+T cells varies in the range of![]() ) [15] [17] . The decline of T is more rapid in the early stage of infection [16] (wherein infected CD4+T cells
) [15] [17] . The decline of T is more rapid in the early stage of infection [16] (wherein infected CD4+T cells ![]() constitute up to 4% of the total number of CD4+T cells T [14] ). The life expectancy of infected CD4+T
constitute up to 4% of the total number of CD4+T cells T [14] ). The life expectancy of infected CD4+T ![]() cells is approximately equal to two days [18] . After an initial period of acute infection, virus increases from below
cells is approximately equal to two days [18] . After an initial period of acute infection, virus increases from below ![]() to
to ![]() or more during the variable course of infection with a sharp increase towards the end of the symptomatic phase [15] . The life span of a virus outside the cell is about 7.2 hours [18] .
or more during the variable course of infection with a sharp increase towards the end of the symptomatic phase [15] . The life span of a virus outside the cell is about 7.2 hours [18] .
5. Conclusion
Mathematical modeling of the dynamics of the development of HIV in the immune system has led to a number of important insights about the dynamics and pathogenesis of HIV infection. A dynamical model of the interaction of HIV and the immune system was presented. We have tried to demonstrate how mathematical modeling can help us to understand HIV pathogenesis. We obtained the solution of the system both analytically and numerically. We applied perturbation and stability theory to obtain our analytic results. Our numerical results are consistent with typical clinical course of HIV infection.