1. Introduction
In the United States, Necrotic enteritis (NE) in broilers is a multi-factorial disease with economic implications of almost $2 billion, annually [1]. The disease is caused by type A strains of Clostridium perfringens (CP) that are specific to poultry with the major toxin type being alpha toxin, and recent evidence suggests the involvement of a novel toxin called NetB [2,3]. CP is ubiquitously found in the environment and is a Gram positive, anaerobic, spore-forming bacteria. Clinical signs of NE include rapid loss in performance, inappetence, severe intestinal damage and are often associated with high mortality [4,5].
Normally, healthy birds harbor a significant number of CP in their intestinal tract. Under specific abnormal conditions, the bacteria are able to colonize and secrete increased amounts of toxins leading to necrosis of the gut mucosa [6,7]. The actual mechanisms of CP pathogenesis are not well understood at this point of time. However, it is widely understood that a coccidial infection is the most common pre-requisite for NE to occur. Damage to the intestinal mucosa is an important factor for CP intestinal colonization and the presence of a coccidial infection is probably the most common causative factor facilitating CP pathogenesis [1,7,8]. Furthermore, there are complex interactions between CP and other members of the gut microflora which are known to be involved in the onset of NE. Additionally, changes in ration, immunosuppression, and withdrawal of the use of anticoccidials or other antimicrobials are also known to predispose birds to NE [1,8].
Several models for NE have been developed in controlled challenge studies in an effort to understand disease progression. Most of these studies involve the use of coccidial challenge as a common predisposing factor in addition to dietary modifications, immunosuppression, and infusion of high CP challenge levels [8-12]. However, recently in a laboratory challenge model study, we demonstrated that neonatal Salmonella typhimurium (ST) infection, followed by an Eimeria and CP challenge caused enhanced development of NE as compared to an Eimeria and CP challenge only [13]. This challenge model appears to be highly reproducible in replicating real world field conditions and most likely integrates all predisposing factors necessary for the onset of NE.
On the other hand, the use of probiotics in agriculture has increased as potential alternatives to antibiotics used as growth promoters [14], and in select cases, for control of specific enteric pathogens [15-19]. For these reasons, the development of effective probiotic products that can be licensed for animal use continues to receive attention [20-22]. Some characteristics are important for the selection of a successful probiotic such as being tolerant to gastrointestinal environment, being able to attach to the intestinal mucosa, and being exclusively competitive with enteric pathogens [23].
During the last 15 years, our laboratories have worked toward the identification of probiotic candidates for poultry which can actually displace Salmonella and other enteric pathogens which have colonized the gastrointestinal tract of chicks and turkeys, indicating that selection of therapeutically efficacious probiotic cultures with marked performance benefits in poultry is possible, and that defined cultures can sometimes provide an attractive alternative to conventional antimicrobial therapy [24,25]. Our studies have been focused on specific pathogen reduction [26-31], performance under commercial conditions [32,33], and effects on both idiopathic [34] and defined enteritis [35,36]. In addition to these comprehendsive studies, preliminary findings from our laboratory indicate that the strains of this probiotic culture exhibits potential probiotic attributes, including the tolerance to pH 3.0, 6.5% of NaCl, high bile salts concentration (0.6%), as well as in vitro antibacterial activity against Salmonella enterica serovar Enteritidis, Escherichia coli (O157:H7), and Campylobacter jejuni [Menconi et al., unpublished data]. Hence, the aim of the present study was to investigate the influence of this Lactobacillus-based probiotic, FloraMax® B-11 (FM-B11) for the control of necrotic enteritis in broiler chickens.
2. Materials and Methods
2.1. Probiotic Culture
FloraMax® B-11 (FM-B11) (Pacific Vet Group USA Inc., Fayetteville AR 72703) is a probiotic culture derived from poultry, consisting of two strains of lactic acid bacterial isolates: Lactobacillus salivarius and Pediococcus parvulus of poultry gastrointestinal origin. Identification has been previously confirmed by 16S rRNA sequence analyses (Microbial ID Inc., Newark, DE 19713, USA) [24].
2.2. In Vitro Assessment of Antimicrobial Activity against Clostridium Perfringens
LAB 18 and LAB 48, the two lab designated strains of FM-B11, were cultured aerobically overnight in Man Rogosa Sharpe (MRS, Catalog no. 288110, Becton Dickinson and Co., Sparks, MD 21152 USA) and screened for in vitro antimicrobial activity against CP. Briefly, ten microliters of lactic acid isolates 18 and 48 of FloraMax®-B11 were placed in the centre of MRS plates. After 24 h of incubation at 37˚C, the plated samples were overlaid with Tryptic Soy Agar (TSA, catalog no. 211822, Becton Dickinson, Sparks, MD) containing 106 cfu/mL of CP and plates were incubated anaerobically. After 24 h of incubation at 37˚C, plates were evaluated for the presence of zones of inhibition.
2.3. Experiment 1, Challenge Organisms
A poultry isolate of Salmonella typhimurium (ST) selected for resistance to nalidixic acid (NA) was used for these trials. An aliquot of ST was thawed and 100 µl of culture was inoculated into 10ml of Tryptic Soy Broth (TSB) (catalog no. 211822, Becton Dickinson, Sparks, MD) and incubated at 37˚C for 24 hours. This was followed by three passages at intervals of eight hours into fresh TSB. Following the last pass, cells were washed with sterile saline (3X) by centrifugation (1864 × g, 4˚C for 15 min). The approximate concentration of ST was estimated spectrophotometrically at 625 nm. In addition, the ST stock solution was serially diluted and plated on Brilliant Green Agar (BGA) (catalog no. 228530, Becton Dickinson, Sparks, MD) plates containing 20 μg/mL NA (catalog no. N4382, Sigma, St Louis, MO) to determine actual concentration.
Eimeria maxima oocysts (EM) were propagated in vivo according to previously published methods [14,15]. A preliminary dose titration study was carried out, offset by 1 week, to determine the Eimeria challenge selection for the present studies. Briefly, broilers at 14 days of age were weighed, divided into three groups and challenged with three different doses of sporulated oocysts of EM by oral gavage. A fourth group of chicks were sham challenged with saline. At 1 wk post-challenge, BW, BWG, and lesion scores were determined. Based on the criterion that the challenge dose caused sub-clinical coccidiosis, a single dose was chosen (Data not shown).
For CP challenge, a strain of CP previously described in a NE challenge model was kindly donated by Dr. Jack. L. McReynolds, USDA-ARS, College Station, TX [17]. A frozen aliquot was shipped on ice to our laboratory and was amplified in TSB with sodium thioglycolate (catalog no. 212081, Becton Dickinson, Sparks, MD). The broth culture was plated on phenyl ethyl alcohol agar (PEA) plates (catalog no. 211539, Becton Dickinson, Sparks, MD) with 5% sheep blood (catalog no. R54012, Remel, Lenexa, KS) to confirm purity, aliquots were made with 25% sterile glycerol and stored at −80°C until further use. A single aliquot was individually amplified in TSB with sodium thioglycolate overnight for challenge studies and the challenge dose was confirmed by plating ten-fold serial dilutions on PEA plates with 5% sheep blood.
2.4. Animal Source
Day-of-hatch, off-sex broiler chickens were obtained from Cobb-Vantress (Siloam Springs, AR, USA) for all the trials mentioned below. All animal handling procedures were in compliance with Institutional Animal Care and Use Committee at the University of Arkansas.
2.5. Experimental Design
Day-of-hatch broiler chicks (n = 141) were randomly allocated to the following treatment groups: 1) Nonchallenged (NC); 2) Challenged (C); 3) Challenged + probiotic (C+ FM-B11). Prior to placement, chicks of groups 2 and 3 received 0.25 mL of ST contained 105 cfu of viable cells by oral gavage. Chicks were placed in 21 sq ft pens on new pine shaving litter with ad libitum access to feed and water. The feed was an unmedicated corn-soy based diet that met National Research Council requirements [25]. At 14, 15 and 16 days of age, all chicks in group 3 were treated with FM-B11 in the drinking water at a concentration of 106 cfu/ml. At 21d of age, all chicks in groups 2 and 3, were individually challenged with 5 × 104 sporulated oocysts of E. maxima by oral gavage. At 26d of age, all chicks in groups 2 and 3, were individually challenged with 108 cfu CP. At 26d of age, body weight (BW) was recorded prior to challenge. The experiment was terminated at 29 days of age and the following parameters were evaluated: NE-associated mortality, CP lesion scores, Ileum CP enumeration by quantitative PCRBW, and body weight gain (BWG). CP lesion scores were evaluated as per Hofacre [22]: 0 = no lesions; 1 = thin-walled and friable intestines; 2 = focal necrosis, gas production and ulceration; 3 = extensive necrosis, hemorrhagic and gas-filled intestines; and 4 = generalized necrosis typical of field cases, marked hemorrhage.
2.6. DNA Isolation and Quantitative PCR for Clostridium Perfringens
Total DNA extraction from ileal samples was achieved using the QIAmp DNA Stool Mini Kit (Qiagen). The manufacturer’s included protocol was modified slightly in the following ways: Ileal contents were removed to include the mucosal layer and diluted 1:5 (w/v) with ice cold PBS + 0.05% Tween 20. One ml of the slurry was added to 1 ml of the included ASL Buffer in a 2.0 ml microcentrifuge tube, vortexed and heated to 70°C for 5 minutes. From this point onwards, the manufacturer’s recommendations were followed to the last step till the DNA was eluted into a final volume of 50 μl. DNA was stored at −20°C until assayed.
Quantitative determination of CP was accomplished using a previously published method with slight modifications [37]. The assay was modified for use on the MX3005P (Agilent Technologies) and Brilliant II QPCR master mix (Agilent Technologies), while all other mixture components, primers, probe and cycling conditions remained as published. A standard curve was prepared using a pure culture of CP serially diluted 10-fold and added to a constant background of ileal content; total DNA isolation was done as previously described.
2.7. Experiment 2, Case Report
Experiment 2 was a unique and remarkable case report of a field outbreak of NE in a commercial broiler farm located in EL Solar, Departamento la Paz, Entre Ríos, Argentina on June 17, 2010. On day 34 of age, the poultry veterinarian diagnosed necrotic enteritis in two of the nine production houses within the complex. The diagnosis was based on clinical history, macroscopic lesions and increased mortality. The control chicken house consisted of 12,000 broiler chickens treated at day 38 with Amoxicillin in the drinking water according to the manufacturer’s specifications (at the time of the outbreak antibiotics were not readily available on site). The second house (Experimental house) involved in the outbreak was treated with FM-B11 in the drinking water for 3 consecutive days (day 34 - 36) according to manufacturer’s instructions. Mortality of the birds in both groups was recorded.
2.8. Statistical Analysis
BW, BWG, ileum log10 CP enumeration and gross NE lesion score data from these experiments were analyzed by ANOVA using the GLM procedure of SAS (© 2008, SAS Institute, Cary, NC), partitioned and treatment means were deemed significant if the P-value was less than or equal to 0.05 (P ≤ 0.05). Mortality data were compared using the chi-square test of independence testing all possible group combinations to determine significance (P ≤ 0.05) for these studies.
3. Results and Discussion
3.1. In Vitro Assessment of Antimicrobial Activity Clostridium Perfringens
Both strains evaluated showed in vitro antibacterial activity against CP (Table 1). The inhibitory activity of LAB against CP has been previously reported and is mainly attributed to the accumulation of primary metabolites such as lactic acid, ethanol, and carbon dioxide and production of other antimicrobial compounds such as bacteriocins [16,29,38-40]. The production levels and proportions among these compounds depend on the biochemical properties of the strains used, and physical and chemical conditions of growth [41-43].
3.2. Body Weight and Body Weight Gain from Experiment 1
In experiment 1, there were no significant (P > 0.05) differences in terms of BW prior to Eimeria challenge on day 21. However, chicks treated with FM-B11 had significantly (P < 0.05) higher BWG after challenge when compared to, control challenge chickens (Table 2).
3.3. Mortality, Lesion Score and Quantification of Clostridium Perfringens from Experiment 1
Total percentage of mortality was evaluated at termina-

Table 1. In vitro assessment of antimicrobial activity of the lactic acid bacteria isolates 18 and 48 present in FloraMax® B-11 against Clostridium perfringens.

Table 2. Effect of Lactobacillus-based probiotic on body weight (BW) and body weight gain (BWG)1 in broilers in a necrotic enteritis model from Experiment 1.
tion of the experiment. Post-mortem analysis was carried out to confirm that mortality was related to necrotic enteritis, which included NE lesion scoring and quantification of CP from ileal content and mucosal scrapings of dead chickens. Total mortality was higher in the C group (46.8%) when compared to C + FM-B11(12.7%) group (Table 3). However, there was no significant (P > 0.05) difference in lesion score between the group C and C + FM-B11 even though, both these groups had significantly (P < 0.05) higher lesions scores than NC. Meanwhile, there was a significant (P < 0.05) reduction in the total number of cfu of CP recovered from the ileal mucosa and content samples in C + FM-B11 when compared to group C (Table 3).
3.4. Mortality from Experiment 2, Case Report
An extraordinary reduction and control of the mortality associated with NE following 3 days of administration of FM-B11 was observed as compared with the control non treated house, where mortality was not controlled until the administration of Amoxicillin at day 38 (Table 4). The results of the present commercial case study are in agreement with several investigators whom have previously reported the ability of lactic acid bacteria to control NE [10,12,44].
4. Conclusion
Probiotics comprising bacteria isolated from poultry sources are increasingly being used in commercial poultry farming [15,19]. The probiotic used in these experiments, FM-B11, contains strains of lactic acid bacteria that were isolated from the gastrointestinal tract of chickens and has consistently proven to exhibit antiSalmonella activity and immunomodulatory effects postchallenge with Salmonella [7,18,33-35]. As previously indicated, a concomitant increase in Salmonella in neonatal chickens, following coccidial challenge may have been controlled by the probiotic preventing further damage. In summary, these results provide evidence which
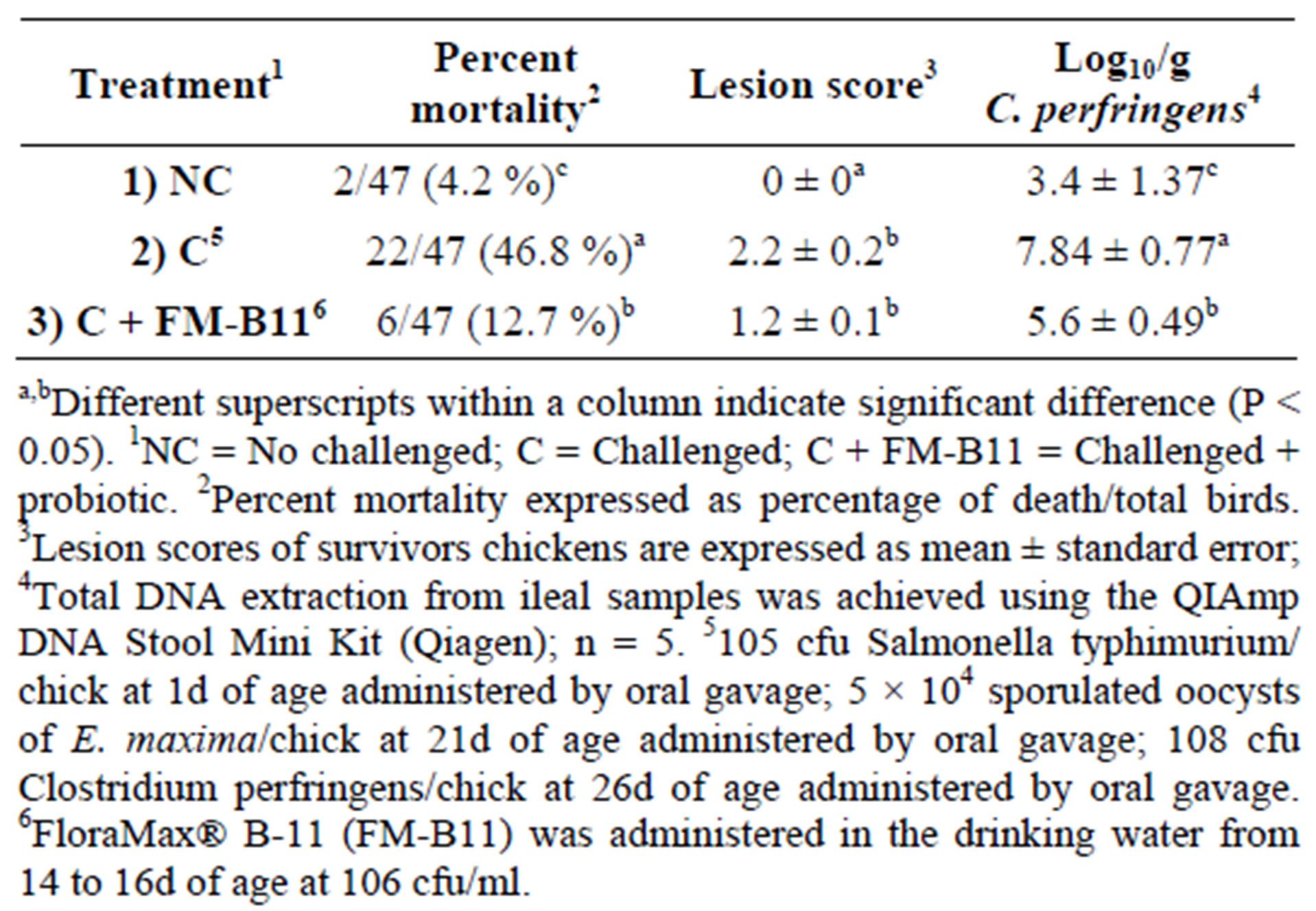
Table 3. Effect of Lactobacillus-based probiotic on necrotic enteritis associated percent mortality and lesion scores in 29 d-old broilers in a necrotic enteritis model from experiment 1.

Table 4. Effect of Lactobacillus-based probiotic on necrotic enteritis associated mortalitya in 34 d-old broilers from Experiment 2, case report.
show that a Lactobacillus-based probiotic, FM-B11, was able to reduce the severities of NE, as a secondary bacterial infection, in a NE laboratory challenge model as well as in a commercial field outbreak of NE.
5. Acknowledgements
The authors are greateful to the Programa de Apoyo a la Superación del Personal Académico (PASPA), Dirección General del Personal Académico (DGAPA) de la Universidad Nacional Autónoma de México (UNAM).
NOTES