World Journal of Cardiovascular Diseases
Vol.4 No.6(2014), Article ID:46491,11 pages DOI:10.4236/wjcd.2014.46040
A Decreased Expression and Functionality of Muscarinic Cholinergic Receptor in Acute Chagas Myocarditis
M. Labrador-Hernández1, L. Jiménez1, G. Leon1, R. López1, W. Mendoza1, C. Bonfante-Rodriguez2, J. Torres-Peraza1,3, R. Bonfante-Cabarcas1*
1Biochemistry Research Unit, School of Medicine, Universidad Centroccidental “Lisandro Alvarado”, Barquisimeto, Venezuela
2Medical Parasitology Research Unit, School of Medicine, Universidad Centroccidental “Lisandro Alvarado”, Barquisimeto, Venezuela
3Cellular Neurobiology and Neurophysiology Laboratories, Applied Medical Research Center, Universidad de Navarra, Pamplona, Spain
Email: *rcabarca@ucla.edu.ve
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


Received 7 April 2014; revised 13 May 2014; accepted 20 May 2014
ABSTRACT
Background: Chagas disease is still a public health problem because of the remaining high prevalence, the expansion of the disease into developed countries and the re-emergences by oral transmission outbreaks. Chagas cardiomyopathy evolves as a consequence of an autonomic unbalance where the parasympathetic tone is undermined. Objective: To determine the functionality and expression of muscarinic cholinergic receptors in acute Chagas disease. Methodology: 62 male, 3-week-old Sprague Dawley rats were assayed; 32 were infected with Trypanosoma cruzi trypomastigotes and 30 were healthy controls. Electrocardiographic studies were conducted in the absence or presence of direct muscarinic (oxotremorine and McN-A-343) or indirect agonists (phenylephrine) or antagonist (pirenzepine). Muscarinic M1 and M2 receptor expression was determined by radioligand [3H]-QNB binding assay and immunoblot. Results: Chagasic acute myocarditis was sustained by electrocardiographic signs and histopathological findings. Bradycardia induced by oxotremorine was significantly higher in healthy rats (HR) and the differences were enhanced by CsCl. In the absence of the agonist, CsCl induced a greater bradycardia in chagasic rats (ChR). In HR McN-A-343 induced tachycardia, however it induces bradycardia in the presence of a acetylcholinesterase inhibitor (neostigmine); no effects were observed in ChR. Pirenzepine induced a higher tachycardia in HR. Phenylephrine in the presence of pirenzepine induced a similar bradycardia in both groups, but recovery was faster in ChR. Muscarinic M1 and M2 receptor density was higher in HR. Conclusion: Muscarinic receptor expression and functionality are decreased in the acute Chagas disease that could impact the evolution and prognosis of the disease.
Keywords:Acute Chagas Disease, Muscarinic Receptors, Oxotremorine, CsCl, McN-A-343, Phenyleprhine, Pirenzepine, [3H]-QNB, Sprague Dawley Rats

1. Introduction
After one hundred years since the discovery of Chagas disease, infection by Trypanosoma cruzi still represents a serious public health problem. Not only in Latin American since nowadays due to the growth of Latin American immigration, it has reached developed countries. The importance of Chagas disease as a public health problem can be summed up in three main reasons: 1) the high residual prevalence in Latin American endemic countries, where the number of infected individuals has been estimated to be 7,694,500 people which represents a prevalence of 1.45%; Bolivia being the southern country with the highest prevalence [1] ; 2) the Latin American migration from endemics areas to non-endemic developed countries such as those in North America, Europe and Australia where the prevalence of these migrant population has been estimated as: 25% in Australia (2005-2006), 9% in Canada (2001), 25% in Spain (2000), 13% in Switzerland, and 0.6% to 5.9% in USA, amongst others. The main concern is blood transfusion related to transmission which has been estimated in Spain in 0.62% [2] -[4] ; 3) Finally, the re-emergence of the disease’s transmission, such as in oral transmission outbreaks where food has been contaminated by triatomines’ dejections; a scenario that has occurred in Brazil, Venezuela and Colombia, among other countries in South America [5] -[10] .
Besides the high mortality of the acute Chagas disease, the most important consequences of Trypanosoma cruzi infection is the subsequent development of chronic chagasic cardiomyopathy, which affects the 20% - 30% of infected patients and is characterized by heart failure, severe arrhythmias, inability to work, extreme poverty and/or sudden deaths [11] . Several hypotheses have been devised to explain the nature of the cardiac dysfunction in Chagas disease. One of them, the neurogenic theory argues that there is a selective damage of parasympathetic intracardiac neurons caused by the mechanic intracellular invasion of the parasite and by the inflammatory process elicited by the infection. Consequently, an autonomic unbalance is displayed, promoting higher sympathetic tone that favors arrhythmias and cardiac remodeling and eventually leading to heart failure [12] [13] .
Destruction of intracardiac parasympathetic neurons during the acute phase of the disease disrupts vagal cholinergic transmission, which should lead to muscarinic receptor up-regulation in postsynaptic membrane effectors’ cells (v.g.: nodal and muscle cells). However from the immunogenic theory’s point of view, muscarinic receptors autoantibodies should cause down-regulation of said muscarinic receptors, because of the agonist like activity of autoantibodies or cell lyses induced by immune response.
We have previously studied muscarinic cholinergic receptors through in vivo and in vitro models of Chagas’ disease [14] [15] . Our results demonstrate that muscarinic receptors were up-regulated in the right ventricular myocardium and peripheral blood mononuclear cells (PBMC) in rats suffering from chronic chagasic myocarditis, and in fetal cardiomyocyte co-culture with T. cruzi/PBMC in non contact fashion or treated with a conditioned medium obtained from T. cruzi-PBMC co-culture [14] . In a different publication, we observed that rats with chronic Chagas disease had parasympathetic disturbances that could be the consequence of alterations on the muscarinic receptor distribution at different neural integration levels [15] . Recently we have also demonstrated a decrease in cardiac response to vagal stimulation in rats with chronic Chagas’ disease, which was reverted by a pharmacological blockage of cholinesterase enzyme with neostigmine [16] .
In the present paper, we studied the functional and molecular expression of muscarinic receptors in Sprague Dawley rats with acute Chagas disease.
2. Materials and Methods
2.1. Animal Sample
We used 62 male 3-week-old Sprague Dawley rats, weighing between 60 and 100 grs that were divided in two groups: healthy (n = 30) and T. cruzi infected (n = 32). The animals part of the T. cruzi infected group were inoculated with 2000 parasites/gr via intraperitoneal of the MHOM/VE/92/2-92-YBM TcI strain; healthy rats were inoculated with the same volume of vehicle (blood). Animals were assayed 3 - 8 weeks after infection; only animals with a provable parasitemia were used. Rats were housed in a number of six in 40 × 25 × 15 cm stainless cages, with free access to food pellets (Ratarina, Protinal, Valencia, Venezuela) and water, 12 hours dark/light cycles and temperature of 25˚C - 28˚C.
2.2. Electrocardiographic Studies
Electrocardiographic recordings were done in a bipolar configuration, with all electrodes located in the subcutaneous tissue, one above the manubrio-sternal joint, the other above xiphoid process and the reference electrode on the abdomen. All electrodes were connected to a BioAmp amplificator (ADInstruments); analog signals were transformed to digital signals by a PowerLab/8sp interphase (ADInstruments), connected to a personal computer using the Chart v4.2.1 software (ADInstruments). Signal capture frequency was set at 400 events/sec and filtered at 60 Hz.
2.3. Pharmacological Protocol
Rats were anesthetized with pentobarbital 40 mg/Kg via ip. Baseline electrocardiographic records were taken thirty minutes after anesthesia was induced. Following that, experimental drugs were administered according to the protocol, each 20 min and the drug effect recorded before next drug administration.
2.4. [3H]-QNB Binding Assay
Myocardial tissue were cut in small pieces, suspended 1:20 w/v in ice cold 1 mM PMSF and homogenized by blending (three strokes of 15-sec bursts with 5-min interval sitting on ice in ultraturrax). The homogenate was filtered through four layers of cotton gauze and centrifuged at 50,000 g for 1 hour, at 4˚C. The supernatant was discharged and the pellet resuspended 1:10 w/v in ice-cold 4 mM EDTA, 0.1 mM PMSF, 25 mM Hepes pH 7.3 buffer. Protein concentration was determined by modified Lowry method.
Receptor density was measured (by quadruplicate) by using saturating concentrations (2 nM) of [3H]-quinuclidinylbenzilate ([3H]-QNB) in equilibrium binding assay conditions according to Perez et al. [17] . Briefly, heart homogenates (500 mg of protein) were incubated with 2 nM [3H]-QNB for 120 min in a final volume of 2.5 ml of 1 mM PMSF in PBS (pH 7.3 at 37˚C). Non-specific binding was defined in the presence of 2 mM atropine. Binding reactions were terminated by vacuum filtration (Brandell, Inc.) through GF/B Whatman glass-fiber filters, washed three times with 5 ml ice-cold 10 mM phosphate buffer solution. Individual filters were placed in glass vials and dried in an air incubator at 60˚C. Once dried, 5 ml of scintillation liquid (23 mM PPO, 0.8 mM POPOP, 10% v/v Triton X-100 dissolved in toluene) was added. Filters were kept in a dark dry room at 20˚C for 12 hours after which radioactivity was measured by liquid scintillation spectrometry (Wallac 1410, Pharmacia, Inc., Finland).
2.5. Immunoblot Analysis
The M1 and M2 muscarinic antibodies, their respective antigenic peptides to whom they were raised and the secondary antibody, all were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Homogenate denatured proteins (60 mg) were fractionated by SDS-polyacrylamide gel electrophoresis (12% polyacrylamide gels) and transferred to nitrocellulose membranes (pore size 0.45 mm; Pierce, Rockford, IL) in a transfer buffer (25 mM Tris-base, 192 mM glycine, 20% methanol, and 0.01% SDS). After the transfer was done, the membrane was washed in Tris Tween-buffered saline (TTBS, 50 mM Tris-base, 159 mM NaCl, 1 mM MgCl2, and 0.05% Tween 20) for 10 min and then incubated in a blocking buffer (BB) containing 1% BSA and 1% casein in TTBS for 24 h at 5˚C, followed by overnight incubation at 4˚C with the primary antibody. The following day, the membrane was washed three times in TTBS and incubated for 1 h with the secondary antibody (bovine anti-goat IgG horseradish peroxidase conjugated) in BB. The membrane was then washed in BB for 15 min. Bound antibodies were detected using chemiluminescent substrate (Luminol Reagent; Santa Cruz Biotech). Negative controls were performed by preincubating the antibodies with the respective peptides against which they are generated.
2.6. Statistical Analysis
All results are presented as mean ± standard error (SEM). We performed a Student’s t test or repeated measure ANOVA followed by Dunnet post test to establish whether there were significant differences between the two groups or the time effect of a drug against the basal conditions respectively, considering statistically significant a p value of <0.05. Analyses were performed using GraphPad Prism 4.
2.7. Ethics
The animals used in this study were manipulated in compliance with APS guiding principles concerning the care and use of laboratory animals, published by the US National Institute of Health and following the experimental animal handling protocol of the Ministry of the Popular Power for Science and Technology (Venezuela).
3. Results
3.1. Basal EKG Records
In Figure 1 typical EKG records are shown. In (a), a normal EKG record from healthy rats is displayed, where a normal wave’s morphology can be seen: amplitude and interval length, as well as a sinusal rhythm. The most frequent disturbances observed in chagasic rats were: disturbances of impulse conductance as AV block; His bundle block and QRS enlargement ((b), (c)); rhythm alterations such as ventricular extrasystoles due to early afterdepolarizations emerging on a prolonged QT interval (d); disturbances of ventricular repolarization as inverted T wave (c) and ventricular hypertrophy ((b), (c)).
3.2. Electrocardiographic Pharmacological Protocols
Oxotremorine 0.1 mg/Kg induced a significant bradycardia at 5 - 25 min and 5 - 20 min in healthy and T. cruzi infected rats, respectively. The bradycardic effect was slightly higher in the healthy rats at all time tested and
 (a)
(a) (b)
(b)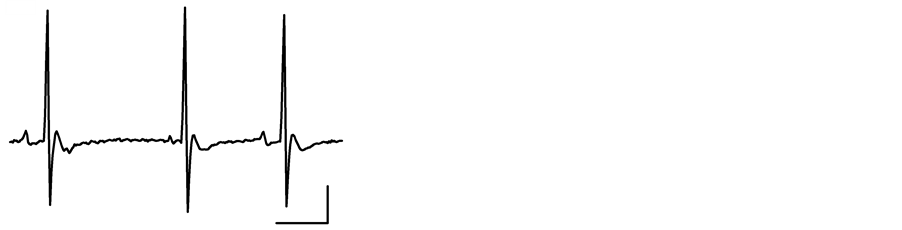 (c)
(c) (d)
(d)
Figure 1. Electrocardiographic disturbances observed in Sprague Dawley rats with acute Chagas disease. Electrocardiographic control basal records were done under pentobarbital (40/Kg) anesthesia in bipolar configuration; analogical signals were captured at 400 Hz in digital format. In (a) is shown a record a healthy rat, while in (b), (c) and (d) are records obtained from different chagasic rats. In (b) conduction disturbances as prolonged QRS interval, His bundle block and repolarization disturbances are shown. In (c) an ischemic and rhythmic disturbances, as well as hypertrophy are displayed. In (d) early afterdepolarizations (arrow) and ventricular extrasystole on T wave are demonstrated. Horizontal bars are equal to 100 msec and vertical bars to 400 µV.
statistically significant at 5 min, when healthy rats were compared against T. cruzi infected rats (Figure 2(a)). On the other hand, 1 mmol/Kg CsCl counteracted the effect of oxotremorine in T. cruzi infected rats’ heart rate but not in healthy rats, this effect allowed to reveal a clear difference between the two groups related to oxotremorine effect (Figure 2(b)). However, when CsCl was applied in the absence of the muscarinic agonist, we observed a paradoxical effect characterized by a significant bradycardia in both groups, but the effect being significantly higher in T. cruzi infected rats (Figure 2(c)).
McN-A-343 a M1 muscarinic receptor agonist increased significantly the heart rate in the healthy rats, but did not have any significant effect in T. cruzi infected rats (Figure 3(a)). In the presence of neostigmine, an acetylcholinesterase inhibitor, McN-A-343 induced a significant bradycardic effect in healthy rats but did not do so in T. cruzi infected rats (Figure 3(b)). To further analyze the pharmacological effect mediated by M1 muscarinic receptors, we explored the effect 0.1 µM pirenzepine, a M1 selective antagonist, observing a time dependant tachycardia in both groups that was significantly higher in healthy rats (Figure 4(a)). Next we explored a vasovagal reflex elicited by phenilephrine in the presence of pirenzepine, obtaining a similar bradycardia but a significant earlier recovery in chagasic rats (Figure 4(b)).
3.3. 3H-QNB Binding Assay
Muscarinic receptor density was decreased in hearts from T. cruzi infected rats when compared to hearts from healthy rats (5.51 ± 0.26 and 8.26 ± 0.35 fmol of [3H]-QNB bound per mg of protein, respectively) (Figure 5(a)).
 (a)
(a) (b)
(b) (c)
(c)
Figure 2. Characterization of bradycardic effect induced by oxotremorine. Rats were anesthetized by pentobarbital 40 mg/Kg vía ip, after 20 - 30 min recordings begin with a control basal record (time 0), after this 0.1 mg/kg oxotremorine (a) or 1 mmol/Kg CsCl (c) were intraperitoneal applied, after 20 min CsCl treated rats were injected with oxotremorine at similar dose (b). Records were made each 5 min. Observe that the bradycardic effect of oxotremorine is higher in healthy (open symbols) when compared with chagasic (fill circles) (a); CsCl increased the differences between two groups about oxotremorine effect (b); CsCl by itself induced bradycardia that was higher in chagasic rats (c). °means p < 0.05 when the drug effect is analyzed against control basal conditions by repeated measure ANOVA, while *means p < 0.05 when healthy and chagasic groups are compared by Student’s t test.
 (a)
(a) (b)
(b)
Figure 3. Pharmacological effect of McN-A-343 in healthy and chagasic rats. Animals were anesthetized by pentobarbital 40 mg/Kg via ip, records start 20 - 30 after anesthesia was induced; McN-A-343 was ip inoculated after control basal was done. In (a) the effect of 0.1 and 1 mg/kg of McN-A-343 is shown, observe that the drug induced tachycardia in healthy rats (open columns) but did not in chagasic ones (fill columns). In (b) the time effect of 1 mg/kg McN-A-343 was assayed in the presence of 0.03 mg/Kg neostigmine, observe that in these conditions McN-A-343 induced bradycardia in healthy rats but still did not have effect in chagasic rats. ° and * means p < 0.05 when the drug effect is analyzed against control basal conditions by repeated measures ANOVA and when healthy and chagasic groups is compared by Student’s t test.
 (a)
(a) (b)
(b)
Figure 4. Pirenzepine effects on basal heart rate and vasovagal response induced by phenylephrine. Animals were anesthetized by pentobarbital 40 mg/Kg via ip, records start 20 - 30 after anesthesia was induced; 0.1 µM pirenzepine was ip inoculated after control basal was done. In a the time effect of pirenzepine is shown, observes that the drug induced tachycardia in both groups, but the increases of heart rate was higher in healthy rats (open circles) when compared with chagasic ones (fill circles). In (b) the time effect of 1 mg/kg phenylephrine was assayed in the presence of pirenzepine, observe that in these conditions both groups had bradycardia, however the recovery to basal heart rate was faster in chagasic rats. ° and * means p < 0.05 when the drug effect is analyzed against control basal conditions by repeated measures ANOVA and when healthy and chagasic groups is compared by Student’s t test.
3.4. Immunoblot
Figure 5(b) and Figure 5(c) show the immunoblot analyses of M1 and M2 muscarinic receptors, where it is shown that the immunogenity against both receptors is higher in the healthy heart homogenates compared with T. cruzi heart homogenate, confirming that muscarinic cholinergic receptors is down regulated in the hearts of T. cruzi infected rats.
4. Discussion
The main finding of the present article is the demonstration that during an acute Chagas’ disease in Sprague Dawley rats there is definitely a muscarinic cholinergic hypofunctionality which was evidenced by electrocardiographic studies, radioligand binding assays and immunoblot; to our knowledge this is the first time that this type of findings are reported.
 (a)
(a) (b)
(b) (c)
(c)
Figure 5. Expression of muscarinic receptors in heart tissue. Total heart tissue homogenates were used for radioligand binding and immune blot assays. Binding assays were done in equilibrium conditions using saturating concentrations of [3H]-QNB (panel a); immunoblot were done using specific M1 and M2 antibodies and revealed by luminol protocol (panel b), densitometric analyses were based on control signals in four different experiments (panel c). Observed that expression of the total muscarinic receptors is lower in chagasic hearts (Tc) when compared with healthy (H) hearts (panel a), this is also true for M1 and M2 muscarinic receptors (b and c panels). * means p < 0.05 when healthy and chagasic groups is compared by Student’s t test.
Firstly, we were able to demonstrate that rats inoculated with T. cruzi developed an acute myocarditis, observing not just the distinctive histopathological findings but also electrocardiographic findings of acute chagasic myocardiopathy (see Figure 1). Recently, our group working on an outbreak of acute Chagas disease in humans observed repolarization disturbances as hallmark signs of acute myocarditis that was reproduced in albino NMRI mice inoculated with the T. cruzi isolates from the outbreak [18] . Likewise, it has been reported that human patients with acute Chagas myocarditis have conduction disturbances, rhythm disorders and ventricular hypertrophia or dilatation [8] [9] .
In the entire anesthetized animals, muscarinic cholinergic hypofunctionality was evidenced as a decreased response to oxotremorine in T. cruzi infected rats, when compared with healthy rats. An effect was potentiated in the presence CsCl. However when CsCl was applied alone, the bradycardic response was significantly higher in T. cruzi infected rats.
CsCl is able to inhibit either K+1 currents activated by acetylcholine (IKACh) or hyperpolarization-activate cation currents (If) in node cells. Both currents are involved in heart’s pacemaker activity and they are regulated by muscarinic receptors [19] . IKACh currents are activated by Gi-coupled M2/M4 muscarinic receptor, where Gi protein directly opens K+1 channels [20] . If is a mixed inward sodium-potassium current that slowly activates on hyperpolarization at voltages in the diastolic range from −60 to −40 mV; also cAMP binds directly to f-channels and increases their open state probability. The cAMP dependence is a particularly relevant physiological property, since it underlies the If—dependent autonomic regulation of heart rate [21] [22] . Muscarinic M2/M4 receptors via Gi-induced adenylciclase inhibition decrease the level of cAMP molecules and shift the If activation range to more negative voltages. This mechanism leads to a decrease of the current at diastolic voltages and therefore to an increase of the flatness of the diastolic depolarization phase and heart rate consequently slows down [23] .
In order to explain the results obtained here, we can speculate that If channels are up regulated while muscarinic M2 receptors are down regulated in chagasic hearts. Because of this, chagasic hearts are more sensitive to CsCl and less sensitive to oxotremorine. Therefore bradycardic response in chagasic rats is closely related to the expression of If channels subunits. It has been reported that an increased number of If subunits and strongly enhanced and accelerated If currents in ventricular hypertrophy in mice have been both related to an up-regulation of HCN1 subunit and to an increased risk of serious arrhythmias [24] [25] .
[3H]-QNB-binding assays sustained a down-regulation of muscarinic receptor in heart chagasic tissue, however, because binding assays were done on washed homogenate tissue, the results could reflect to a certain extent the binding properties of inflamed tissue, including inflammatory cells, degenerated cardiomyocites and interstitial edema which is known to have lesser binding capacity and lesser expression of muscarinic receptor than healthy cardiomyocytes themselves. Conversely, Peraza et al., 2008 [14] have demonstrated an up-regulation of muscarinic receptors expressed in cardiomyocyte in an in vitro model of Chagas disease, where cardiomyocyte did not directly contact T. cruzi, but they were in contact with the inflammatory cytokines produced by the interaction between T. cruzi and PBMC. Taken together, both results indicate that heart cell lysis by either T. cruzi invasion or by the inflammatory process account for a heart muscarinic down-regulation observed in acute Chagas myocarditis. Meanwhile, an up-regulation induced by cytokines should occur once the acute process is solved by the immunological system and cardiac tissue remodeling occurs in the subacute and chronic phases of the Chagas disease.
Immunoblot data sustained binding data and also suggested a M1 receptor down-regulation, which is in agreement with McN-A-343 electrocardiographic data, where we were able to observe that McN-A-343 failed to induce a tachycardia in T. cruzi infected rats. McN-A-343 is a selective M1 muscarinic agonist that stimulates muscarinic transmission in sympathetic ganglia. This explains the tachycardic effect of the drug [26] .
In the presence of an acetylcholinesterase inhibitor, McN-A-343 induced a significant bradycardia instead of tachycardia in healthy rats, having no effect in T. cruzi infected rats. McN-A-343 is a partial agonist with similar affinity at all five muscarinic acetylcholine receptor subtypes and its relative selectivity depends on a higher efficacy at the M1 (and M4) subtypes [26] . Being a partial agonist, its action is also dependent on other factors, such as receptor density and coupling efficacy between receptor activation and tissue response [26] . In addition, McN-A-343 is a bitopic agonist, because it binds to an allosteric site on the M2 receptor as well as to the orthosteric site [27] . Bradycardic effect observed in healthy rats could be explained by a reciprocal potentiation agonist effect of endogenous acetylcholine and McN-A-343 on M2 muscarinic receptors, given that inhibition of acetylcholinesterase by neostigmine increases acetylcholine concentration in the synaptic cleft. In T. cruzi infected rats because of the down-regulation of the muscarinic receptor, the bradycardic effect was not observed.
Pirenzepine caused a significantly higher tachycardia in healthy rats. Pirenzepine is a selective M1 muscarinic antagonist with a ki value in the order of 3.16 - 15.9 nM and also interacts with M2 muscarinic receptors with ki of 0.2 - 0.5 µM [28] . Consequently, the effects at a doses of 0.1 µmol/Kg could be given to M2 receptor blocking effects, which are evident because of the higher densities of M2 muscarinic receptor in heart tissue. A smaller response in T. cruzi infected rats further confirms a muscarinic receptor down-regulation.
Phenylephrine has been widely used to assay vasovagal reflex and the overall integrity of vagal efferent innervations of the heart. Here in the presence of pirenzepine, the recovery from the bradycardia induced by phenylephrine was faster in T. cruzi infected rats. This could imply that acetylcholine reserves in synaptic terminals at sinoatrial nodes were lower as a consequences of vagal denervation caused by parasite invasion of the neurons, degeneration caused by periganglionic inflammation or antineuronal autoimmune reaction [29] or that, conversely, sympathetic tone is higher in chagasic rats [30] .
Our results are consistent with a decreased cholinergic function during acute chagasic myocarditis. Decreased cholinergic tone during the course of a heart disease could account for homeostatic disturbances responsible for heart failure’s poor prognosis [31] [32] . Cholinergic tone protects heart against ischemic and inflammatory phenomena. When cholinergic system is affected, adrenergic tone predominates leading to hemodynamic and remodeling changes responsible for heart failure. This autonomic dysregulation has long been recognized as a mediator of increased mortality and morbidity in myocardial infarction and heart failure [33] . Sympathovagal imbalance can lead to increased heart rate, excess release of proinflammatory cytokines, dysregulation of nitric oxide (NO) pathways, and arrythmogenesis [33] . Wang et al. (2003) [34] have demonstrated that the vagus nerve can inhibit significantly and rapidly the release of macrophage TNF, and attenuate systemic inflammatory responses, and has termed it the “cholinergic anti-inflammatory pathway”. The essential macrophage acetylcholine-mediated (cholinergic) receptor that responds to vagus nerve signals has been identified as the nicotinic acetylcholine receptor α7 subunit that is required for acetylcholine inhibition of macrophage TNF release. Recent studies have suggested a strong association between cardiac arrhythmias and local inflammation that potentiate reciprocally a high sympathetic tone [33] .
Our results support the hypothesis that cholinergic dysfunction in Chagas disease is the result in part of muscarinic receptor down-regulation and hipofuncionality. The importance of our findings is to have been able to demonstrate that alterations in the cardiac muscarinic cholinergic transmission are present in the acute Chagas disease. This physiopathological phenomenon could impact the evolution and prognosis of the disease. Since the inhibitory and protective effect of the cholinergic system is decreased, favoring cardiac remodeling induced by excitatory and inflammatory mediators (for example norepinephrine and angiotensin), cardiac muscarinic cholinergic functionality could be useful as prognostic marker of the disease.
Acknowledgments
This paper was funded by CDCHT (Consejo de Desarrollo Cientifico, Humanístico y Tecnológico) at Universidad Centroccidental “Lisandro Alvarado”, Barquisimeto, Venezuela; Projects’ Number 006-ME-2008.
References
- Guhl, F. and Lazdins-Helds, J.K. (2007) Grupo de trabajo científico. Reporte sobre la enfermedad de Chagas. OMS, Electronic Adress: http://whqlibdoc.who.int/hq/2007/TDR_SWG_09_spa.pdf.
- Schmunis, G. (2007) Epidemiology of Chagas Disease in Non-Endemic Countries: The Role of International Migration. Memórias do Instituto Oswaldo Cruz, 102, 75-85. http://dx.doi.org/10.1590/S0074-02762007005000093
- Jackson, Y., Gétaz, L., Wolff, H., Holst, M., Mauris, A., Tardin, A., Sztajzel, J., Besse,V., Loutan, L., Gaspoz, J.M., Jannin, J., Vinas, P.A., Luquetti, A. and Chappuis, F. (2010) Prevalence, Clinical Staging and Risk for Blood-Borne Transmission of Chagas Disease among Latin American Migrants in Geneva, Switzerland. PLOS Neglected Tropical Diseases, 4, e592. http://dx.doi.org/10.1371/journal.pntd.0000592
- Piron, M., Vergés, M., Muñoz, J., Casamitjana, N., Sanz, S., Maymó, R.M., Hernández, J.M., Puig, L., Portús, M., Gascón, J. and Sauleda, S. (2008) Seroprevalence of Trypanosoma Cruzi Infection in At-Risk Blood Donors in Catalonia (Spain). Transfusion, 48, 1862-1868. http://dx.doi.org/10.1111/j.1537-2995.2008.01789.x
- Toso, M.A., Vial, U.F. and Galanti, N. (2011) Oral Transmission of Chagas’ Disease. Revista Medica de Chile, 139, 258-266.
- de Noya, B.A., Díaz-Bello, Z., Colmenares, C., Ruiz-Guevara, R., Mauriello, L., Zavala-Jaspe, R., Suarez, J.A., Abate, T., Naranjo, L., Paiva, M., Rivas, L., Castro, J., Márques, J., Mendoza, I., Acquatella, H., Torres, J. and Noya, O. (2010) Large Urban Outbreak of Orally Acquired Acute Chagas Disease at a School in Caracas, Venezuela. The Journal of Infectious Diseases, 201, 1308-1315. http://dx.doi.org/10.1086/651608
- Benchimol-Barbosa, P.R. (2010) Trends on Acute Chagas’ Disease Transmitted by Oral Route in Brazil: Steady Increase in New Cases and a Concealed Residual Fluctuation. International Journal of Cardiology, 145, 494-496. http://dx.doi.org/10.1016/j.ijcard.2009.08.030
- Cutrim, F.S., Almeida, I.A., Eda, G.G. and Silva, A.R. (2010) Chagas’ Disease in the State of Maranhão, Brazil: Record of Acute Cases from 1994 to 2008. Revista da Sociedade Brasileira de Medicina Tropical, 43, 705-708. http://dx.doi.org/10.1590/S0037-86822010000600021
- Pinto, A.Y., Valente, S.A., Vda, C.V., Ferreira Jr., A.G. and Coura, J.R. (2008) Acute phase of Chagas Disease in the Brazilian Amazon Region: Study of 233 Cases from Pará, Amapá and Maranhão Observed between 1988 and 2005. Revista da Sociedade Brasileira de Medicina Tropical, 41, 602-614.
- Nicholls, R.S., Cucunubá, Z.M., Knudson, A., Flórez, A.C., Montilla, M., Puerta, C.J. and Pavía, P.X. (2007) Acute Chagas Disease in Colombia: A Rarely Suspected Disease. Report of 10 Cases Presented during the 2002-2005 Period. Biomédica, 27, 8-17.
- Teixeira, A.R., Hecht, M.M., Guimaro, M.C., Sousa, A.O. and Nitz, N. (2011) Pathogenesis of Chagas’ Disease: Parasite Persistence and Autoimmunity. Clinical Microbiology Reviews, 24, 592-630. http://dx.doi.org/10.1128/CMR.00063-10
- Dávila, D.F., Donis, J.H., Torres, A. and Ferrer, J.A. (2004) A Modified and Unifying Neurogenic Hypothesis Can Explain the Natural History of Chronic Chagas Heart Disease. International Journal of Cardiology, 96, 191-195. http://dx.doi.org/10.1016/j.ijcard.2003.06.015
- Da Cunha, A.B. (2003) Chagas’ Disease and the Involvement of the Autonomic Nervous System. Revista Portuguesa de Cardiologia, 22, 813-824.
- Peraza-Cruces, K., Gutiérrez-Guédez, L., Perozo, D.C., Lankford, C.R., Rodríguez-Bonfante, C. and Bonfante-Cabarcas, R. (2008) Trypanosoma Cruzi Infection Induces Up-Regulation of Cardiac Muscarinic Acetylcholine Receptors in Vivo and in Vitro. Brazilian Journal of Medical and Biological Research, 41, 796-803. http://dx.doi.org/10.1590/S0100-879X2008000900009
- Labrador-Hernández, M., Suárez-Graterol, O., Romero-Contreras, U., Rumenoff, L., Rodríguez-Bonfante, C. and Bonfante-Cabarcas, R. (2008) The Cholinergic System in Cyclophosphamide-Induced Chagas Dilated Myocardiopathy in Trypanosoma-Cruzi-Infected Rats: An Electrocardiographic Study. Investigación Clínica, 49, 207-224.
- Bonfante-Cabarcas, R., Hincapié, E.L., Hernández, E.J., Zambrano, R.F., Mancini, L.F., Mena, M.D. and RodríguezBonfante, C. (2013) Electrophysiological and Pharmacological Evaluation of the Nicotinic Cholinergic System in Chagasic Rats. BMC Pharmacology and Toxicology, 14, 2. http://www.biomedcentral.com/2050-6511/14/2
- Pérez, C.C., Tobar, I.D., Jiménez, E., Castañeda, D., Rivero, M.B., Concepción, J.L., Chiurillo, M.A. and Bonfante-Cabarcas, R. (2006) Kinetic and Molecular Evidences That Human Cardiac Muscle Express Non-M2 Muscarinic Receptor Subtypes that Are Able to Interact Themselves. Pharmacological Research, 54, 345-355. http://dx.doi.org/10.1016/j.phrs.2006.07.001
- Alvarado-Tapias, E., Miranda-Pacheco, R., Rodríguez-Bonfante, C.C., Velásquez, G., Loyo, J., Gil-Oviedo, M., Mogollón, N., Pérez-Aguilar, M.C., Recchimuzzi, G., Espinosa, R., Carrasco, H.J., Concepción, J.L. and Bonfante-Cabarcas, R.A. (2012) Electrocardiographic Repolarization Abnormalities Are Characteristic Signs of Acute Chagasic Cardiomyopathy. Investigación Clínica, 53, 378-394.
- Van Borren, M.M., Verkerk, A.O., Wilders, R., Hajji, N., Zegers, J.G., Bourier, J., Tan, H.L., Verheijck, E.E., Peters, S.L., Alewijnse, A.E. and Ravesloot, J.H. (2010) Effects of Muscarinic Receptor Stimulation on Ca2+ Transient, cAMP Production and Pacemaker Frequency of Rabbit Sinoatrial Node Cells. Basic Research in Cardiology, 105, 73-87. http://dx.doi.org/10.1007/s00395-009-0048-9
- Tamargo, J., Caballero, R., Gómez, R., Valenzuela, C. and Delpón, E. (2004) Pharmacology of Cardiac Potassium Channels. Cardiovascular Research, 62, 9-33. http://dx.doi.org/10.1016/j.cardiores.2003.12.026
- Bucchi, A., Barbuti, A., Difrancesco, D. and Baruscotti, M. (2012) Funny Current and Cardiac Rhythm: Insights from HCN Knockout and Transgenic Mouse Models. Frontiers in Physiology, 3, 240. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3387723/
- Scicchitano, P., Carbonara, S., Ricci, G., Mandurino, C., Locorotondo, M., Bulzis, G., Gesualdo, M., Zito, A., Carbonara, R., Dentamaro, I., Riccioni, G. and Ciccone, M.M. (2012) HCN Channels and Heart Rate. Molecules, 17, 4225-4235. http://dx.doi.org/10.3390/molecules17044225
- DiFrancesco, D. and Tromba, C. (1988) Muscarinic Control of the Hyperpolarization-Activated Current (If) in Rabbit Sino-Atrial Node Myocytes. The Journal of Physiology, 405, 493-510.
- Fernández-Velasco, M., Goren, N., Benito, G., Blanco-Rivero, J., Boscá, L. and Delgado, C. (2003) Regional Distribution of Hyperpolarization-Activated Current (If) and Hyperpolarization-Activated Cyclic Nucleotide-Gated Channel mRNA Expression in Ventricular Cells from Control and Hypertrophied Rat Hearts. The Journal of Physiology, 553, 395-405. http://dx.doi.org/10.1113/jphysiol.2003.041954
- Hofmann, F., Fabritz, L., Stieber, J., Schmitt, J., Kirchhof, P., Ludwig, A. and Herrmann, S. (2012) Ventricular HCN Channels Decrease the Repolarization Reserve in the Hypertrophic Heart. Cardiovascular Research, 95, 317-326. http://dx.doi.org/10.1093/cvr/cvs184
- Mitchelson, F.J. (2012) The Pharmacology of McN-A-343. Pharmacology & Therapeutics, 135, 216-245. http://dx.doi.org/10.1016/j.pharmthera.2012.05.008
- Valant, C., Gregory, K.J., Hall, N.E., Scammells, P.J., Lew, M.J., Sexton, P.M. and Christopoulos, A. (2008) A Novel Mechanism of G Protein-Coupled Receptor Functional Selectivity. Muscarinic Partial Agonist McN-A-343 as a Bitopic Orthosteric/Allosteric Ligand. The Journal of Biological Chemistry, 283, 29312-29321. http://dx.doi.org/10.1074/jbc.M803801200
- Caulfield, M.P. and Birdsall, N.J. (1998) International Union of Pharmacology. XVII. Classification of Muscarinic Acetylcholine Receptors. Pharmacological Reviews, 50, 279-290.
- Marin-Neto, J.A., Cunha-Neto, E., Maciel, B.C. and Simões, M.V. (2007) Pathogenesis of Chronic Chagas Heart Disease. Circulation, 115, 1109-1123. http://dx.doi.org/10.1161/CIRCULATIONAHA.106.624296
- Torres, A., Dávila, D.F., Gottberg, C.F., Donis, J.H., de Bellabarba, G.A. and Ramoni-Perazzi, P. (2000) Heart Rate Responses to a Muscarinic Agonist in Rats with Experimentally Induced Acute and Subacute Chagasic Myocarditis. Revista do Instituto de Medicina Tropical de São Paulo, 42, 219-224. http://dx.doi.org/10.1590/S0036-46652000000400007
- Sabbah, H.N. (2011) Electrical Vagus Nerve Stimulation for the Treatment of Chronic Heart Failure. Cleveland Clinic Journal of Medicine, 78, S24-S29. http://dx.doi.org/10.3949/ccjm.78.s1.04
- Schwartz, P.J. (2011) Vagal Stimulation for Heart Diseases: From animals to Men—An Example of Translational Cardiology. Circulation Journal, 75, 20-27. http://dx.doi.org/10.1253/circj.CJ-10-1019
- Das, U.N. (2011) Vagal Nerve Stimulation in Prevention and Management of Coronary Heart Disease. World Journal of Cardiology, 3, 105-110. http://dx.doi.org/10.4330/wjc.v3.i4.105
- Wang, H., Yu, M., Ochani, M., Amella, C.A., Tanovic, M., Susarla, S., Li, J.H., Wang, H., Yang, H., Ulloa, L., Al-Abed, Y., Czura, C.J. and Tracey, K.J. (2003) Nicotinic Acetylcholine Receptor Alpha7 Subunit Is an Essential Regulator of Inflammation. Nature, 421, 384-388. http://dx.doi.org/10.1038/nature01339
Abbreviation List
AV: Atrioventricular BB: Blocking buffer cAMP: Cyclic adenosine monophosphate ChR: Chagasic rats ECG: Electrocardiogram EDTA: Ethylenediaminetetraacetic acid HCN: Hyperpolarization-activated cyclic nucleotide-gated channels.
[3H]-QNB: [3H]-quinuclidinyl benzilate.
HEPES: hydroxyethyl piperazineethanesulfonic acid.
HR: Healthy rats.
Ip: intraperitoneal McN-A-343: 4-(m-chlorophenyl-carbamoyloxy)-2-butynyltrimethylammonium chloride)
PBMC: Peripheral blood mononucleated cell PMSF: Phenylmethylsulfonyl fluoride POPOP: Di(phenyl-5-oxazolyl-2)-1, 4-benzene PPO: 2, 5-Diphenyloxazole SDS: Sodium dodecyl sulphate T. cruzi: Trypanosoma cruzi TTBS: Tris Tween-buffered saline
NOTES

*Corresponding author.