Advances in Microbiology
Vol.4 No.4(2014), Article ID:43988,5 pages DOI:10.4236/aim.2014.44026
Behavior of the Fungus Colletotrichum gloeosporioides (Penz & Sacc.), Which Causes Bitter Rot in Apples after Harvesting
Sideney Becker Onofre, Dirlane Antoniazzi
Departamento de Ciências Biológicas e Farmacêuticas, Faculdade Educacional de Francisco Beltrão—FEFB, União de Ensino do Sudoeste do Paraná—UNISEP, Francisco Beltrão, Paraná, Brasil
Email: becker@unisep.edu.br, beckerside@gmail.com
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


Received 7 February 2014; revised 7 March 2014; accepted 14 March 2014
ABSTRACT
The apple is the second most important fruit in Brazil. However, apple cultivars are susceptible to several diseases that can cause losses after harvesting. Bitter rot is caused by the fungus Colletotrichum gloeosporioides and is one of the most damaging summer diseases. The goal of this work was to evaluate the behavior of this fungus in four apple cultivars grown in Brazil (Fuji, Gala, Golden and Green) under two treatments: direct inoculation and isolated fungus. The fungus was isolated by taking fragments from infected fruits, which were stored on Potato Dextrose Agar (PDA) in a laboratory. For the direct treatment, the fungus was removed from infected fruits and directly inoculated into healthy fruits. After inoculation, the fruits were kept at room temperature and the halos of degradation were evaluated every 48 hours. The results demonstrated that direct inoculation was more pathogenic, that the Gala cultivar was the most resistant to the pathogen, and that the Golden cultivar was the most susceptible.
Keywords:Diseases; Biotechnology; Phytopathogens; Fungi; Pathology

1. Introduction
The southern region of Brazil is a prominent producer of apples, 85% of which are consumed in natura as an indispensable source of vitamins and minerals for humans [1] . In 2002, the state of Santa Catarina produced 857 tons of apples, which was 55.3% of the national production.
During and after harvesting, the fruits are prone to infection by several pathogens because the resistance in their peels and pulp decreases during maturation. During disbranching, injuries to the fruits are an essential factor in the establishment of diseases, once the fruits become more susceptible to fungi. According to Bleicher [2] , production losses of apples in southern Brazil can reach 50% just from bitter rot.
According to Tavares [3] , post-harvest loss of tropical fruits in Brazil is about 30% for commercialized products. The most common diseases caused by fungi are responsible for 80% - 90% of the total loss caused by phytopathogens.
Bitter rot is one of the major diseases of post-harvested fruits, and is mainly caused by Colletotrichum gloeosporioides. The symptoms of this disease are round, large, necrotic lesions. The edges of the lesions appear slightly more elevated than the center, where the masses of conidia are produced. As a result, the fruits rot; however, the symptoms of the infection vary among the different hosts.
This pathogen can affect leaves at any stage of development, new branches, flowers and fruits. During the rainy season, frizzing of new branches is the most common symptom; the branches become purple, then dark brown, dry and fragile [4] .
This pathogen is important in the degradation of fruits, where it can stay latent (e.g., in immature fruits) until conditions become suitable for its development, which generally takes place when the fruit ripens. In high humidity, fresh fruits and vegetables are prone to attack by phytopathogenic fungi and bacteria, as well as to physiological degradation during the period between harvesting and consumption [5] .
The colonies of Colletotrichum gloeosporioides vary from light to dark grey and the conidia are salmon colored. Their development is favored by relatively high temperature and humidity, reduced ventilation and illumination, and the presence of injuries on the fruits. The conidia are released and spread in many ways, such as when the acervuli are humid, by raindrops, wind, and insects [3] .
Tavares [3] states that the conidia germinate in the presence of water and then produce appresoria, which initiate the process of penetrating the host’s tissue. At first, the hyphae grow both interand intracellularly, but produce few or no symptoms in the tissues. According to this author, the fungus survives in the petioles and old leaves, and is spread by rain or wind to flowers and young fruits where it remains latent until the fruits ripen.
When flower buds are infected, the rot is exhibited as a dark region from the peduncle to part or all of the fruit. In ripe fruits, the infection generally begins after harvesting, which causes small, depressed, light brown lesions. These become deeper and soft and are usually covered by orange tufts of conidia, and this often results in fruit rot [4] .
According to Pio-Ribeiro and Mariano [6] , in ripe fruits the pulp under the dark spots rots, causing the fruits to fall. When the conditions of development of the disease are favorable, the signs of the pathogen are easily visible in the affected areas because concentric punctuations are formed by the fruiting bodies of the fungus [7] .
Presently, because the use of synthetic fungicides is restricted both nationally and internationally, research is being conducted on alternative means of controlling this rot after harvesting. Genetic improvements in the development of cultivars more resistant to diseases have significantly advanced, which has diminished the use of fungicides by more than 80%. The effects of these types of advances are immediate, not only because they decrease production costs, but also because they protect the producer, the health of the consumers and preserve the environment.
Producers and researchers have found that it is possible to produce quality fruit, and reduce the use of agrochemicals and environmental impact, when cultivation practices are used in an integrated manner [8] . One of the purposes of integrated production is to manage the cultivar so that the plants can express their natural resistance to plagues and pathogens, which helps protect useful organisms.
This work evaluated the behavior of the fungus Colletotrichum gloeosporioides (Penz), which causes bitter rot, in stored fruits of four apple cultivars.
2. Materials and Methods
2.1. Isolation of the Fungus
This research was carried out at the laboratory of Microbiology of Unisep—União de Ensino do Sudoeste do Paraná—Unit Campus Francisco Beltrão, from April to July, 2013. The isolates were obtained from lesions of apple fruits (Malus domestica) from cultivars grown in Palmas, PR, with symptoms characteristic of the disease caused by Colletotrichum gloeosporioides.
The isolation was carried out by removing fragments from the sites with lesions. These fragments were disinfected in 1% sodium hypochlorite solution for two minutes, then placed in 40% alcohol for one minute and then washed with sterilized distilled water. After disinfection, the fragments were dried on sterilized filter paper and transferred to Petri dishes containing Potato Dextrose Agar (PDA) medium.
To grow the fungus, the temperature was kept at 26˚C for eight days. The fungus was then identified using a microculture technique, taxonomic keys, and other literature.
2.2. Behavior of the Fungus
To setup the experiment, the fruits were washed with soap in running water. They were then hydrothermally treated, to prevent and control pathogens that cause post-harvesting diseases, by immersing them in hot water (50˚C) for five minutes. Following this, the fruits were disinfected by immersing them in a 1% sodium hypochlorite solution for 10 minutes and then washed in sterilized distilled water before the inoculations were made.
The cultivars evaluated were Fuji, Gala, Golden and Green, and the inoculations were made in two ways: 1) direct inoculation and 2) isolated fungus. In the treatment with the isolated fungus, the fruits were perforated with a metal punch that produced holes 7 mm in diameter and 3 mm deep. A disc of the culture medium containing the mycelium of C. gloeosporioides, which was the same size, was then placed in the hole. In the direct treatment, the fungus was removed from a fruit that showed symptoms of the disease and inoculated directly into healthy fruits through holes made with the punch. After inoculation, the fruits were kept at room temperature to evaluate the development of the disease. Evaluations of the halos of degradation were made every 48 hours and the diameters of the lesions were measured at right angles with a millimeter ruler. The evaluations ended after 336 hours, when the fruits had advanced lesions.
3. Results and Discussion
The results, obtained by observing the behavior of C. gloeosporioides exposed to four apple cultivars under two conditions (isolation and direct inoculation), are summarized in Figures 1 and 2.
The analysis of the data shows that the pathogenicity of the fungus at 96 hours was higher, because it was more aggressive, when the inoculation was made with the fungus that was isolated and stored in a laboratory in culture medium. After 96 hours, this behavior changed and greater pathogenicity was seen in the samples where the inoculation was transferred directly from contaminated fruits to healthy fruits. These fruits had larger inhibition halos than those observed on fruits inoculated with the stored isolate, which can be observed at 144 to 336 hours of evaluation.
The analysis also verified that the behavioral data are significant, because in the first hours of observation
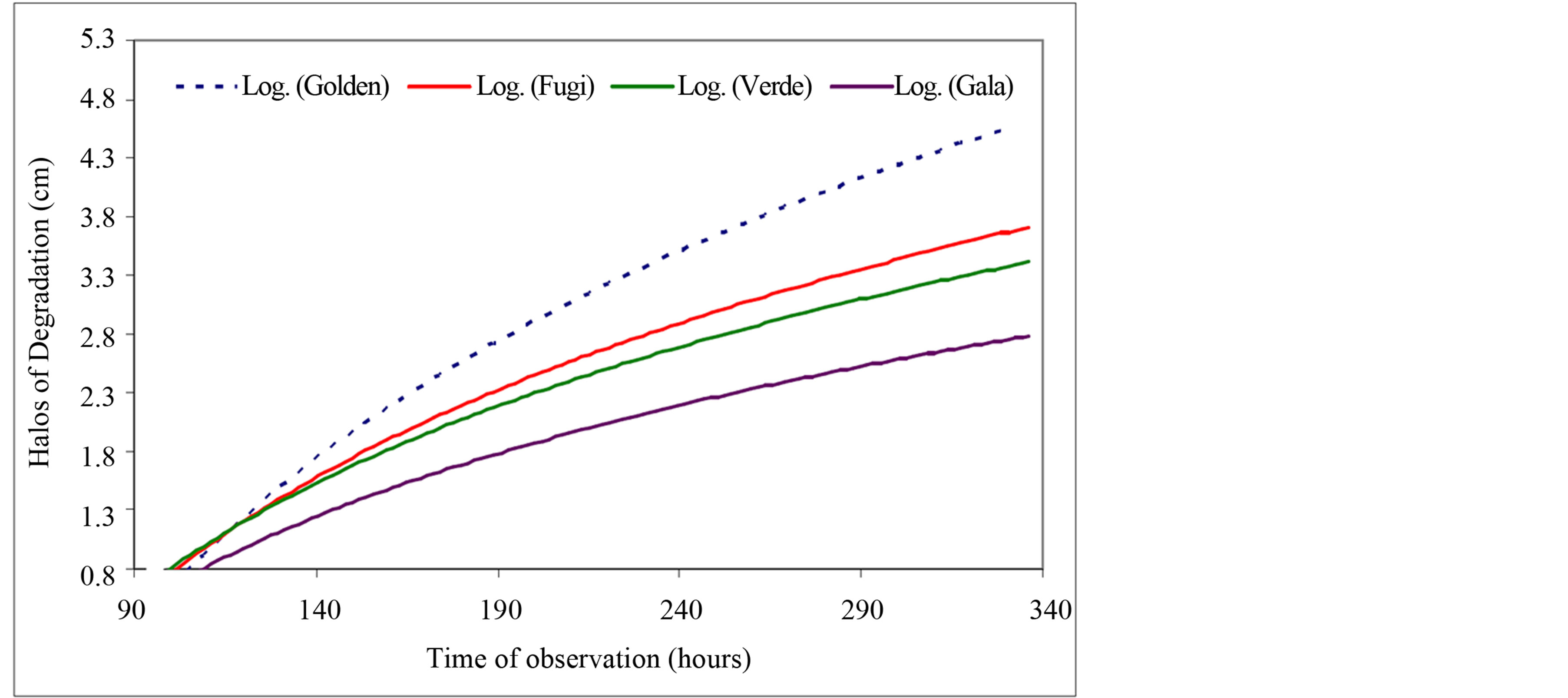
Figure 1. Kinetics of degradation caused by the fungus Colletotrichum gloeosporioides on four apple cultivars (Fuji, Gala, Golden and Green) exposed by direct inoculation.

Figure 2. of the degradation caused by the fungus Colletotrichum gloeosporioides on four apple cultivars (Fuji, Gala, Golden and Green) exposed to the stored isolate.
(also called Lag Phase) the fungi were adapting; the microorganisms were beginning to express their factors of pathogenicity, especially the enzymes and extracellular proteins that act on the fruit tissues.
The data refer to the final observation time (336 hours) provide an overview of the general and final behavior of the fruit disease during storage and ripening because direct inoculation was statistically greater (at the level of 5%) than the treatment that used the isolated fungus. The Gala cultivar was the most resistant to the pathogen, and had halos of degradation that were 2.87 cm in the direct treatment and 1.42 cm in the treatment with the isolate. The Golden cultivar was the most susceptible to the fungus, with halos of degradation that were 4.88 cm (direct treatment) and 3.65 cm (isolated fungus).
This behavior exhibited by the fungus could be related to the inhibition of some genes linked to the pathogenicity of the microorganisms, the synthetic, non-natural culture medium the fungus was stored in, or to changes in the metabolism of the fungus caused by the low storage temperature.
The data obtained in this work corroborate those observed by Camilo [9] , who reported that the injuries caused to the epidermis of apples facilitate the entrance, establishment and development of bitter rot.
Silva [10] , working with the same fungus under similar conditions, verified that this fungus caused the degradation of stored fruits. This author worked with mango, papaya, passion fruit and guava and the data recorded matches those of this work, because they demonstrated that all the isolates showed pathogenicity in all fruit species evaluated. Guava is the least susceptible host to many isolates of C. gloeosporioides. The most susceptible fruits are papaya and the mango cultivars Tommy Atkins and Rose, where lesions are known to grow faster compared to those on other fruits.
Muniz et al. [11] suggest the occurrence of groups of pathogenic specialization and that the different pathogenicities of the isolates could be the consequence of environmental interactions or the genotype of the hosts. These authors verified that isolates of C. gloeosporioides from different hosts (avocado, mangabeira, passion fruit and acerola) are pathogenic to cashew, which indicates that cultures of C. gloeosporioides from different hosts can serve as a source of inoculate and could have implications in the epidemiology and control of the disease.
According to Freeman et al. [12] , the differentiation of Colletotrichum species based on the host is not an accurate method to identify fungi in this genus, because taxa such as C. gloeosporioides, C. dematium, C. acutatum, C. graminicola infect a wide range of plants. As verified by Dianese et al. [13] , isolates of C. gloeosporioides from Eucalyptus pellita infected guava, mango, papaya, red pepper and E. grandis, but did not infect orange and eggplant.
The data obtained in this work agree with the results of Peres et al. [14] , who found that Colletotrichum gloeosporioides can infect avocado, apple, ponkan and peach. These authors reported that the isolates of this fungus have the potential to affect several tropical fruits through crossed infections, as observed in this study when the isolates from all the fruits showed aggressiveness in all of the hosts; although, at different degrees [15] [16] .
4. Conclusions
Based on the results, the direct inoculate had a higher degree of pathogenicity than the treatment exposed to the isolated fungus stored in culture medium.
The inoculated fungus caused the disease in all the cultivars studied. Halos of degradation grew the slowest on the Gala cultivar, which was the most resistant to Colletotrichum gloeosporioides. The Golden cultivar showed the largest halos of degradation and was considered the most susceptible to the disease.
Conflict of Interest
The authors declare no conflict of interest.
References
- Bleicher, J. (1980) Fatores predisponentes para a ocorrência da podridão amarga, Glomerella cingulata, em maça. Florianópolis, Empresa Catarinense de Pesquisa Agropecuária, 17p. Comunicado Técnico, 36.
- Bleicher, J. (2002) História da Macieira. In: EPAGRI. A cultura da Macieira. Florianópolis: Empresa de Pesquisa e Extensão Rural de Santa Catarina—EPAGRI, Vol. 11, pp. 29-35.
- Tavares, G.M. (2004) Controle químico e hidrotérmico da antracnose em frutos de mamoeiro na pós-colheita. 55p. Dissertação (Mestrado em Fitopatologia)—Universidade Federal de Lavras, Lavras-MG.
- Junqueira, N.T.V. (2000) Doenças e pragas. Porto Alegre-RS: Cinco Continentes, 225-247.
- Zambolim, L. (2002) Controle integrado de doenças em pós-colheita de frutíferas tropicais. In: Simpósio de controle de doenças de plantas: patologia pós-colheita de frutas e hortaliças, 2., 2002, Lavras-MG. Anais... Lavras: UFLA/FAEP, 139-145.
- Pio-Ribeiro, G. and Mariano, R.L. (1997) Doenças do maracujazeiro. Manual de Fitopatologia. 3 ed. 2, Agronômica, Ceres, São Paulo.
- Carneiro, S.M.T.P.G., Pignoni, E., Vasconcellos, M.E.C. and Gomes, J.C. (2007) Eficácia de extratos de Nim para o controle de oídio do feijoeiro. Summa Phytopathologica, 33, 34-39. http://dx.doi.org/10.1590/S0100-54052007000100005
- Fachinello, J.C. (2003) Situação e perspectiva da produção integrada na Europa. In: Produção Integrada de Frutas: o caso da maça no Brasil. Bento Gonçalves: EMBRAPA Uva e Vinho, 21-33.
- Camilo, A.P. (1989) Genetic Resistance in Malus to Glomerella cingulata (Stoneman) Spaulding & Von Schrenk: Sorces of Resistance, Leaf Infection, Progeny Evaluation, and Pathogenecity. 116f. Tese, Cornell University, Ithaca, USA.
- Silva, K.S. (2005) Avaliação da agressividade de Colletotrichum gloeosporioides (Penz) em manga, mamão, maracujá e goiaba na pós-colheita. Vitória da Conquista—BA: UESB, 50p. (Dissertação—Mestrado em Agronomia, Área de Concentração em Fitotecnia).
- Muniz, M.F.S., Santos, R.C.R. and Barbosa, G.V.S. (1998) Pathogenicity of Colletotrichum gloeosporioides Isolates on Some Tropical Fruits. Summa Phytopathologica, 24, 177-179.
- Freeman, S., Katan, T. and Shabi, E. (1998) Characterization of Colletotrichum Species Resposible for Anthracnose Diseases of Various Fruits. Plant Disease, 82, 596-605. http://dx.doi.org/10.1094/PDIS.1998.82.6.596
- Dianese, J.C., Ribeiro, W.R.C. and Moraes, T.C.A. (1984) Colletotrichum gloeosporioides (Penz.) Sacc, em lesões de ramos de Eucalyptus pellita afetado pelo Mal do Rio Doce. Fitopatologia Brasileira, 9, 426-429.
- Peres, N.A.R., Kuramae, E.E., Dias, M.S.C. and Souza, N.L. (2002) Identification and Characterization of Colletotrichum spp. Affecting Fruit after Harvest in Brazil. Journal of Phytopathology, 15, 128-134. http://dx.doi.org/10.1046/j.1439-0434.2002.00732.x
- Oliveira, J.A. (1991) Efeito do tratamento fungicida em sementes e no controle de tombamento de plântulas de pepino (Cucumis sativas L.) e pimentão (Capsicum annanum L.). 117p. Dissertação (Mestrado em Fitossanidade)— Universidade Federal de Lavras, Lavras-MG.
- Sales Junior, R. and Costa, F.M. (2004) Utilização de Azoxistrobina no Controle da Antracnose da Mangueira. Fitopatologia Brasileira, 29, 193-196. http://dx.doi.org/10.1590/S0100-41582004000200012