N. SHIOMI, M. AKO
308
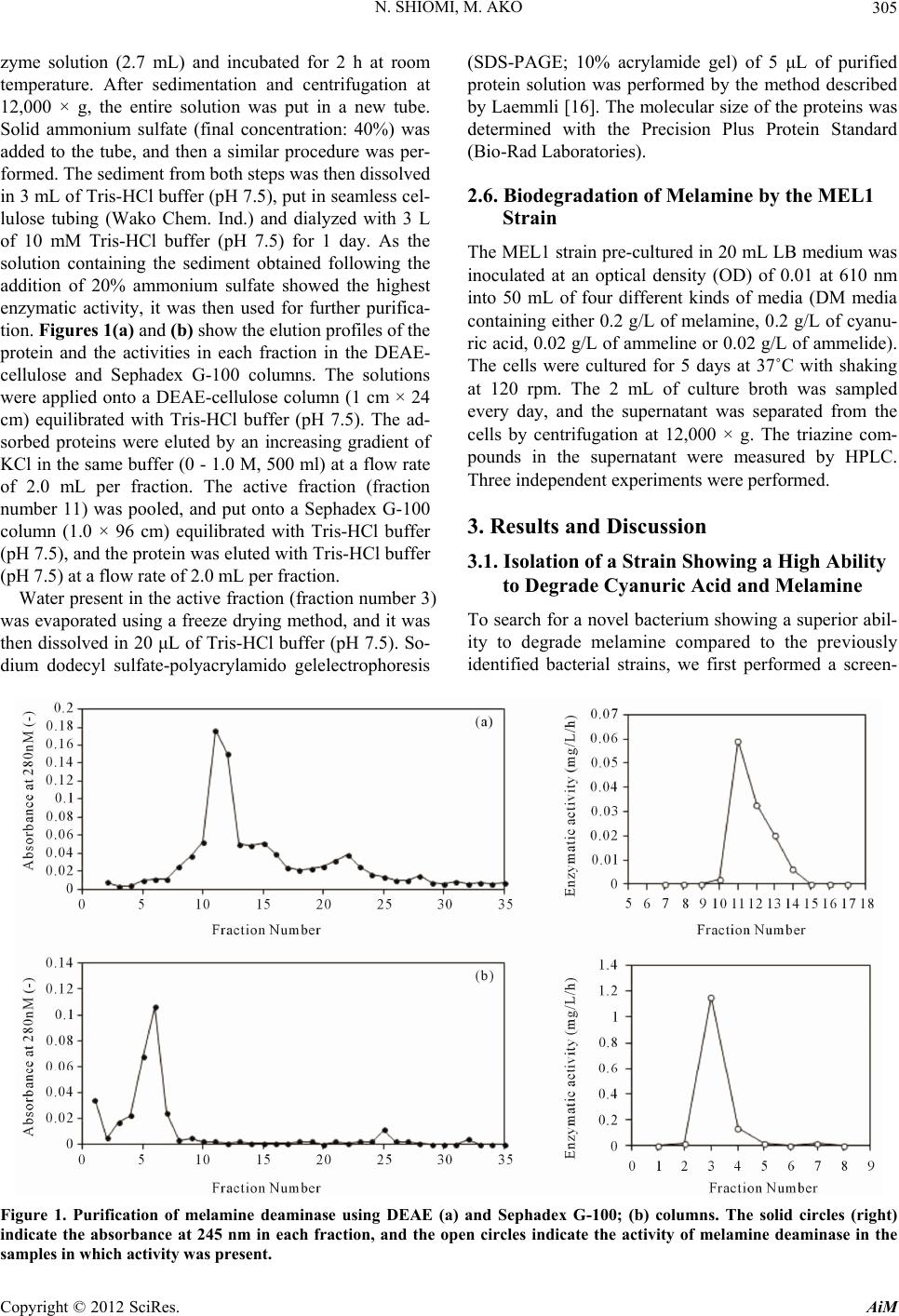
Figure 5. The time courses of the changes in the melamine
concentration and the concentrations of its byproduct when
the MEL1 strain was cultured in the DM medium contain
n. Melamine was generally
egraded to ammeline, ammelide and cyanuric acid in
tu
ning melamine, as in Figure 4. As shown
in
Sasaguri and
ble assistance in conducting
B JAPAN Corporation for
pact
,
Vol. 13, No. 40, 2008, pp. 1-2.
[2] C. A. Brown, enga, B. Puschner,
0
-
ing melamine (as shown in Figure 4). Bars, means ± SD of 3
independent experiments.
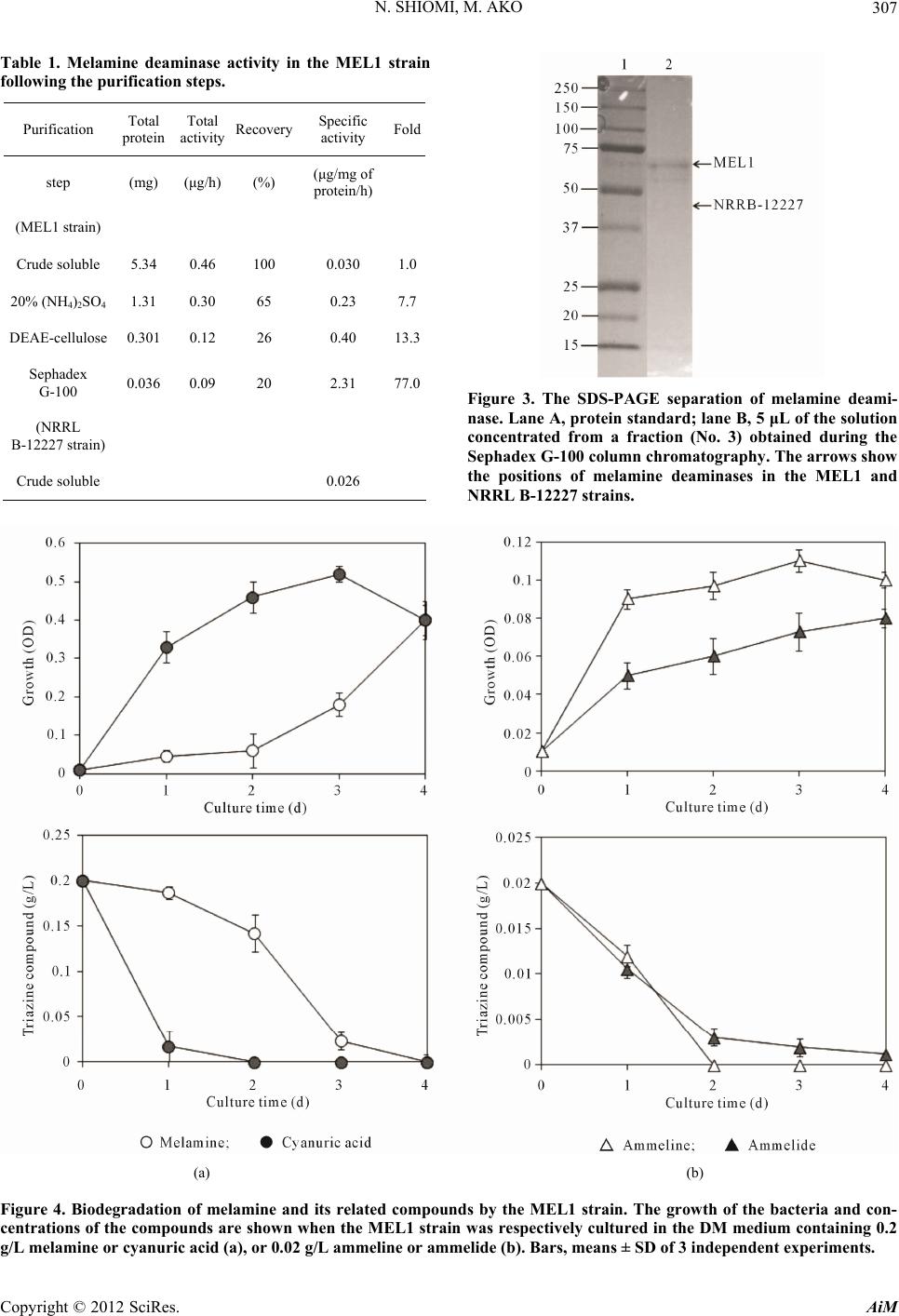
Finally, we examined the characteristics of melamine
degradation in the MEL1 strai
d
rn in bacteria [18]. Figures 4(a) and (b) show the
growth of the bacteria and the concentration of triazine
compounds when the MEL1 strain was respectively cul-
tured in the DM media containing melamine, cyanuric
acid, ammeline or ammelide. Lower concentrations were
used in case of ammeline and ammelide, because they
were difficult to dissolve at higher concentrations. The
MEL1 strain could degrade all four of the triazine com-
pounds as sole nitrogen sources, and the rates of mela-
mine and cyanuric acid degradation were as high as those
of other melamine-degrading strains [9-12]. Moreover,
the degradation rate for cyanuric acid was faster than that
for melamine.
Figure 5 shows the concentrations of melamine and its
by-products when the bacteria were cultured in the DM
medium contai
Figure 5, cyanuric acid, ammeline and ammelide were
not detected during the degradation of melamine. Be-
cause of toxicity, the minimal accumulation of cyanuric
acid during the biodegradation of melamine is preferred,
and therefore, the MEL1 strain might be useful for the
bioremediation of melamine.
4. Acknowledgements
Our group is grateful to Ms. Syoko Mrs.
Madoka Yasui for their valua
our research, and the NCIM
the identification of the MEL1 strain. This research was
REFERENCES
[1] D. Coulimbier, C. Hepner, S. Fabiansson, A. Tarantola, A.
Cochet, P. Kreidl and R. Reintjes, “Melamine Contami-
nation of Dairy Products in China-Public Health Im
on Citizens of the European Union,” Eurosurvieillance
supported by grants-in-aids from the Central Laboratory
and the School of Human Sciences at Kobe College.
K.-S. Jeong, R. H. Popp
D. M. Miller, A. E. Ellis, K.-I. Kang, S. Sum, A. M. Cis-
tola and S. A. Brown, “Outbreaks of Renal Failure Asso-
ciated with Melamine and Cyanuric Acid in Dogs and
Cats in 2004 and 2007,” Journal of Veterinary Diagnostic
Investigation, Vol. 19, No. 5, 2007, pp. 525-531.
doi:10.1177/10406387070190051
[3] S. A. Tittlemier, B. P. Lau, C. Ménard, C. Corrigan, M.
Sparling, D. Gaertner, K. Pepper and M. Feeley, “Mela-
mine in Infant Formula Sold in Canada: Occurrence and
Risk Assessment,” Journal of Agricultural Food Chemis-
try, Vol. 57, No. 12, 2009, pp. 5340-5344.
doi:10.1021/jf9005609
[4] W. C. Anderson, S. B. Turnipseed, C. M. Karbiwnyk, S.
B. Clark, M. R. Madson, C. M. Gieseker, R. A. Miller, N.
G. Rummel and R. Reimschuessel, “Determination and
Confirmation of Melamine Residues in Catfish, Trout,
Tilapia, Salmon, and Shrimp by Liquid Chromatography
with Tandem Mass Spectrometry,” Journal o
f Agricul-
tural Food Chemistry, Vol. 56, No. 12, 2008, pp. 4340-
4347. doi:10.1021/jf800295z
[5] S. L. Rovner, “Anatomy of a Pet Food Catastrophe”,
Chemical & Engineering News, Vol. 86, No. 18, 2008, pp.
41-43. doi:10.1021/cen-v086n019.p041
[6] D. Park, T. K. Kim, Y. J. Choi, S. H. Lee, D.-K. Bae, G.
Yang, Y.-H. Yang, S. S. Joo, E.-K. Chioi, B. Ahn, J.-C.
Kim, K.-S. Kim and Y.-B. Kim, “Increased Nephrotoxic-
ity after Combined Administration of Melamine and
Cyanuric Acid in Rats,” Laboratory Animal Research,
Vol. 27, No. 1, 2011, pp. 25-28.
doi:10.5625/lar.2011.27.1.25
[7] Y. Qin, X. Lv, J. Li, G. Qi, Q. Diao, G. Liu, M. Xue, J.
Wang, J. Tong, L. Zhang and K. Zhang, “Assessment of
Melamine Contamination in Crop, Soil and Water in
China and Risks of Melamine Accumulation in Animal
Tissues and Products,” Environment Internarional, Vol.
36, No. 5, 2010, pp. 446-452.
doi:10.1016/j.envint.2010.03.006
[8] D. R. Shelton, J. S. Karns, G. W. Mccarty and D. R.
Durham, “Metabolism of Melamine by Klebsiella ter-
ragena,” Applied and Environmental Microbiology, Vol.
63, No. 7, 1997, pp. 2832-2835.
[9] K. Takagi, K. Fujii, K. Yamaza
saki, “Biodegradation of Melamin
ki, N. Harada and A. Iwa-
e and Its Hydroxy De-
d Properties of an s-Tri-
rivatives by a Bacterial Consortium Containing a Novel
Nocarduoides Species,” Applied Microbiology and Bio-
technology, Vol. 94, No. 6, 2012, pp. 1647-1656.
[10] J. K. Karns, “Gene Sequence an
Copyright © 2012 SciRes. AiM