Open Journal of Ecology
Vol.4 No.9(2014), Article
ID:46618,11
pages
DOI:10.4236/oje.2014.49043
Competitive Consumption of the Lake Kinneret (Israel) Plankton by Hypophthalmichthys molitrix and Sarotherodon galilaeus
Moshe Gophen
Migal-Research Institute in the Galilee, Kiryat Shmone, Israel
Email: Gophen@Migal.org.il
Copyright © 2014 by author and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/
![]()
![]()

Received 5 April 2014; revised 5 May 2014; accepted 13 May 2014
ABSTRACT
An outdoor tank experiment was carried out for the assessing of the impacts of Sarotherodon galilaeus (Galilee Saint Peter’s Fish; Cichlidae) (SG) and Hypophthalmichthys molitriox (Silver Carp; Cyprinidae) (SC) on Lake Kinneret (Israel) plankton. In order to evaluate the interaction effect, the experiments were of replicated 2 × 2 factorial design: TG × SC. A lot of interaction effects were indicated showing that the effects of the two fishes were not independent and potentially competitors. SG suppressed most crustaceans and rotifers while increasing gross and net primary production and chlorophyll concentration. SC had less intense effects on zooplankton than SG. Although SC suppressed most crustaceans and rotifers, it had less interaction effects than SG. SC had no statistically significant effects on phytoplankton production or chlorophyll concentration. It is suggested that these experiments indicate that although the plankton community impacts of SG and SC do differ, both fishes utilize similar food resources in Lake Kinneret. Fingerlings of SG and SC are planted in Lake Kinneret annually aimed at the improvement of fishermen’s income and prevention of water quality deterioration. SC is known as efficient consumer of Microcystis. It is therefore recommended to limit SC introduction to periods when Microcystis is abundant.
Keywords:S. galilaeus, H. molitrix, Lake Kinneret, Plankton Consumption

1. Introduction
The silver carp (SC) is known as a filter feeder, which has a specialized apparatus capable of filtering small particles. The gill rakers are fused into a sponge-like filter, and an epibranchial organ secretes mucus which adhere trapped particles. Strong buccal pump forces are operated for the implementation of high quantity of water flow through the filter apparatus. The major food sources consumed by SC are known to be the phytoplankton but they also consume zooplankton and detritus. Due to their phytoplanktivory trait, they are successfully used for controlling Cyanobacteria.
Since early 1990s, the Kinneret ecosystem has undergone changes in limnology [1] -[4] . These included increase in P, decline in N and the ensuing decrease of the TN/TP mass ratio that showed N limitation in the epilimnion. The practical implication of that is a threat to water quality, when Cyanobacteria blooms increase. P increase and N decrease create N limitation for Peridinium growth and optimum for Cyanobacteria [1] [2] . New condition of N limitation in Lake Kinneret was found to be optimal for the growth of Cyanbacteria, diatoms and chlorophytes. A change of the phytoplankton composition was therefore documented, especially decline in Peridinium but increase of the other phytoplankton. These ecosystem modifications initiated change in the food resources and consequently in feeding habits of the lake fish. To ensure reasonable water quality, it is important to maintain the high grazing pressure of zooplankton on nano-phytoplnkton. Therefore, removal of the unwanted zooplanktivorous bleaks by fishery management [2] [3] and introduction of SC, an efficient consumer of Microcystis, might be beneficial. These changes could culminate in deterioration of water quality but can be probably confounded by stocking of SC. This exotic carp cannot reproduce in Lake Kinneret and its introduction might have an advantage because the fish is a well-known consumer of the toxic cyanophyte Microcystis [5] [6] and can partially compensate for the fishermen’s income during low seasons for the S. galilaeus (SG) fishery. SC has been stocked into Lake Kinneret since 1969 and during 2000-2009 introduction is averaged as (Sanovsky, pers. commun.) follows: Total Introduction −0.29 × 106/year, lake population size −0.01 × 106/year and mean density −1.7 Kg/ha. During the 1970-1980’s, management programs intended to improve water quality in Lake Kinneret have proposed to reduce Peridinium densities by increasing populations of SG, a commercially valuable native cichlid that feed heavily on Peridinium [7] [8] . Nevertheless, due to the food web structure modifications during the 2000’s, there is a concern about potentially competing between stocked cichlids and SC and potential adverse impacts on the plankton community structure. A study on the impacts of SG and SC on the lake Kinneret plankton community was carried out. SC is native to eastern Asia and has been widely introduced throughout the world mostly as a productive pond fish. Both SG and SC consume plankton as filter-feeders but their relative impacts on Lak Kinneret plankton populations have not been studied previously.
2. Methods
Two 5 m3 outdoors mesocosm experiments were conducted to asses and compare the impacts of SG and SC on the Kinneret plankton populations. SG was collected from the lake and SC from fish rearing ponds. Prior to experimentation, fish were acclimated for 7 days in 5m3 outdoor tanks continuously supplied with water and plankton pumped from the lake and supplemented with commercial fish food. The experiments were conducted in a linear array of 8 cylindrical 5 m3 black plastic tanks (1.9 m diameter). Experiments were of 2 × 2 factorial design (presence and absence of SG × presence and absence of SC) allowing us to examine main and “interaction effects” of each fish species. Two replicates of each treatment combination (no fish, SG, SC, and SG + SC) were assigned to tanks using a randomized block assign. Blocks consisted of tanks 1 through 4, and tanks 5 through 8. Two independent 12 - day duration experiments were performed. These experiments were conducted when Peridinium were still abundant in the lake but dominance was rapidly shifting to nano-phytoplankton. The combined species treatments (SG + SC) had approximately doubled the total biomass in comparison with single species treatment. This double biomass was necessary for interaction effects which assess whether the effects of the fish species were independent. This design holds the impact of each fish species constant within the context of the factorial design, and the equivalent density and biomass of each fish species in the presence or absence of the other holds intra-specific competition while allowing inter-specific to vary. Although the fish biomass densities used in our experiments exceed those likely to be observed for these species in Lake Kinneret, they were selected to allow assessment of community responses during relatively shortterm experiments while minimizing tank effects. The community responses measured is qualitatively similar to impacts expected from these fishes in the lake though they may differ in magnitude. It is also recognized that confinement of the fish to the tank system must have reduced their option for behavioral flexibility and may also affect the outcome of such experiments. Nevertheless, this approach is valuable as one of several possible experimental techniques for the study of fish impacts on plankton community structure.
Four fish of each species per tank repetition were individually weighed and measured before and after experiments. Average total fish biomass for SG added to each experimental tank during first and second experiment respectively was 421 g (range 398 - 428 g) and 436 g (range 427 - 446 g). The total biomass of four specimen of SC used in the first and second experiments were 428 (range 410 - 505 g), and 453 g (range 379 - 577 g), respectively, which is correspond to values between “Median” and “High” presented by Xia Zhang et al. [9] . No fish mortality or change in fish biomass was observed during these experiments. Tanks were filled at the beginning of each experiment with water pumped from app. 30 meters offshore at a depth of 1.5 m. No water were added to the tanks after initial filling. After filling the tanks we supplemented zooplkankton in them with plankton collected 1 - 2 km offshore. Sixteen 10-meter vertical hauls using a 300-µm-mesh, 44-cmdiameter conical plankton net were pooled and equal portions of the collected material added to each tank, to assure the presence of large bodied limnetic zooplankton in all tanks.
Tanks were mixed for 2 hours daily by an air-lift mixer system. Air from blower was injected 1 m below the water surface into 5-cm-diameter plastic pipes suspended from floats in each tank. Mixers moved a water volume equivalent to tank volume in approximately 1 hour, de-stratifying and aerating the tanks. Because Peridinium divide primarily at night and are sensitive to agitation during this daytime mixers were operated only during the day. Tanks were sampled initially and at 3 day intervals for 12 days. Mean tank water temperatures ranged from 26.4˚C to 30.4˚C during the experiments. On each sampling date, a single composite water sample was produced for each tank by mixing 5 replicate samples collected with a 2.5-m long, 1.5 cm-diameter plastic pipe lowered to within 10 cm of the tank bottom. Aliquots were taken for analysis of chlorophyll and particle size distribution. Total chlorophyll measurement was done on un-filtered water and nanoplankton chlorophyll determination on 25 µm mesh filtered water [10] on triplicates of each sample. Samples for determination of particle size frequency distribution were preserved (Lugol’s Iodine solution) and counted using Coulter Particle Counter (Model ZB). Measurements of primary production were performed on all sampling dates using light-dark-bottles-oxygen production method. Zooplankton samples were collected from each tank at the conclusion of the mixing period by a single vertical haul of a 16-cm diameter, 63 µm mesh net. At least 200 organisms or one third the total sample volume from each tank was counted using dissecting microscope. Indirect fish effect on filter-feeding zooplankton production was studied by placing Ceriodaphnia spp. in protected small enclosures inside the big tanks. These small enclosures were made of 15 cm long plexiglass tube 5 cm in diameter with two ends blocked by 45 µm mesh netting. Ten adult Ceriodaphnia spp. were placed in each chamber at the beginning of each experiment and 4 chambers were suspended at mid-water in the center of each tank. During the first experiment chambers were suspended only in fishless and those containing SG tanks. In the second experiment chambers were placed in all tanks. Data were analyzed using multivariate analysis of repeated measures [11] . This procedure sums data across sampling dates into a uni-variate test to detect treatment effects. Statistical significance was inferred at the level of p < 0.1.
3. Results
Both fish species significantly reduced densities of most zooplankton species during each experiment (Figures 1-4). Evaluation of SG and SC main effects and interactions that are presented in Tables 1-4 were achieved by comparing the mean values for treatments containing the species SG and both for SG and SC and Both for SC with those from which it is absent, SC and None for SG and SG and None for SC as done separately for each experiment. SG suppressed Ceriodaphnia spp., Bosmina, spp, Mesocyclops sp., cyclopoid copepodids, cyclopoid nauplii, Collotheca sp. Polyarthra sp., Keratella spp., Trichocerca sp., and Brachionus spp., in the two experiments. SC impacts on zooplankton differed slightly between experiments. In the first experiment SC significantly suppressed Ceriodaphnia spp., Bosmina spp., Mesocyclops sp., cyclopoid copepodids, cyclopoid nauplii, Collotheca, sp., Polyarthra sp., Keratella spp., and Trichocerca sp., while in the second experiment they suppressed Ceriodaphnia spp., Mesocyclops sp., cyclopoid copepodids, Keratella spp., and Trichocerca sp. Significant interaction effects were detected for most zooplankton taxa in one or both experiments. These indicate that the impacts of a fish species on the plankton community were significantly affected by the presence of the other species in the Tank. Interactions occurred between the two fish species and basically their impacts were not additives. It is probably because they are slightly differs in food items selection. Significant fish effect on chlorophyll concentration were no detected with multi-variate profile analysis, although examination of data over the course of the
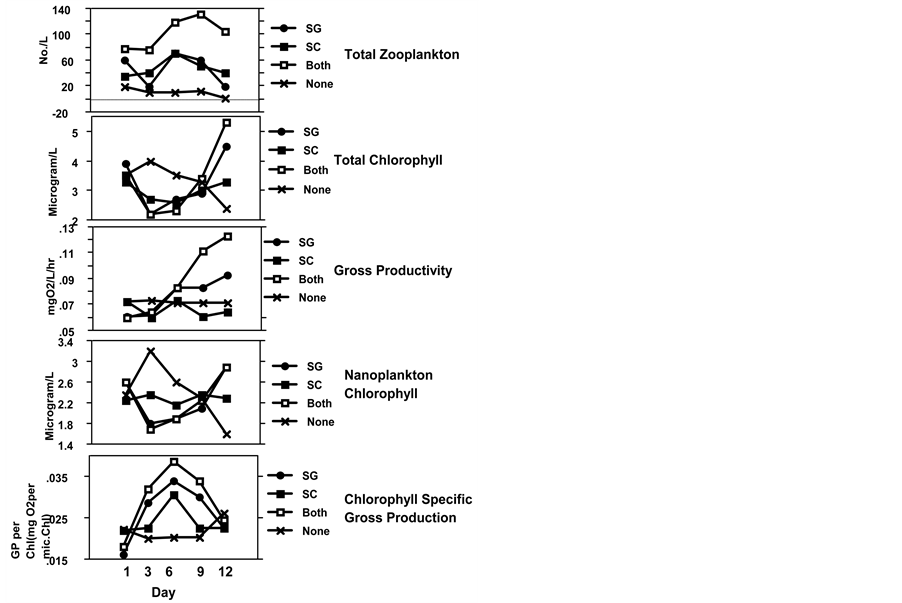
Figure 1. Treatment mean values averaged for two experiments, (SD’s ranged between 5% - 60%) during the 12th days of each experiment. Parameters presented include: Total Zooplankton abundance (No/L); Total Chlorophyll concentration (µg/L); Total Nano-plankton Chlorophyll concentration (µg/L; Gross Primary Productivity (mg O2/L/hr); and Chlorophyll Specific Gross Primary Production (mg O2 produced per µg Chlorophyll per hr). Errata: In upper panel “Both” is “None “ and “None” is “Both”.
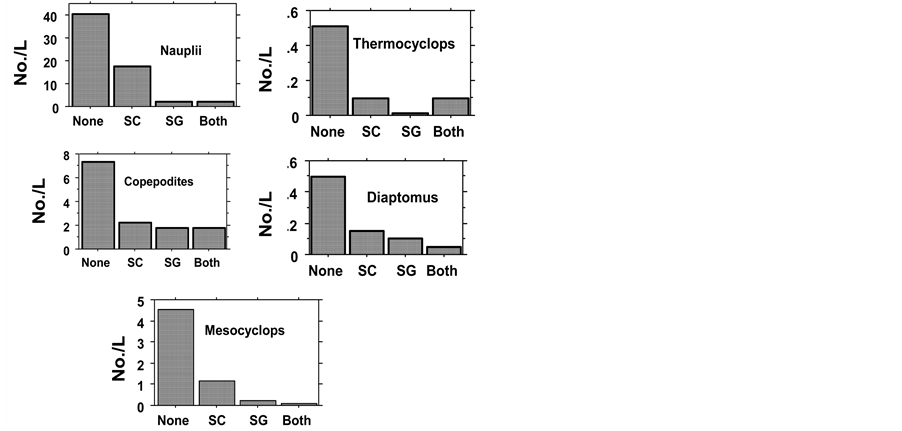
Figure 2. Mean (two experiments all samples) of Copepoda in Fishless (None), H. molitrix (SC), S. galilaeus (SG), SC + Sc (Both). Probability values are given in Table3
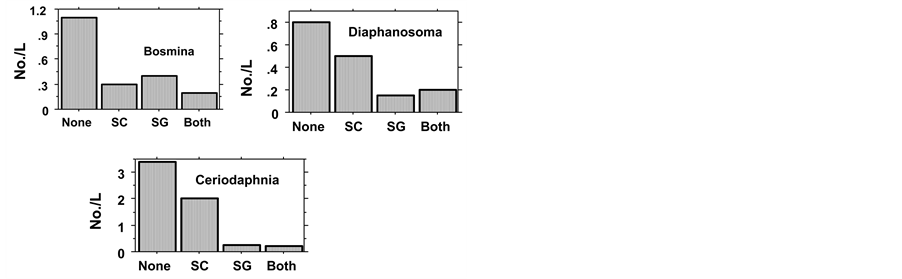
Figure 3. Mean (two experiments all samples) of Cladocera in Fishless (None), H. molitrix (SC), S. galilaeus (SG), SC + Sc (Both). Probability values are given in Table3
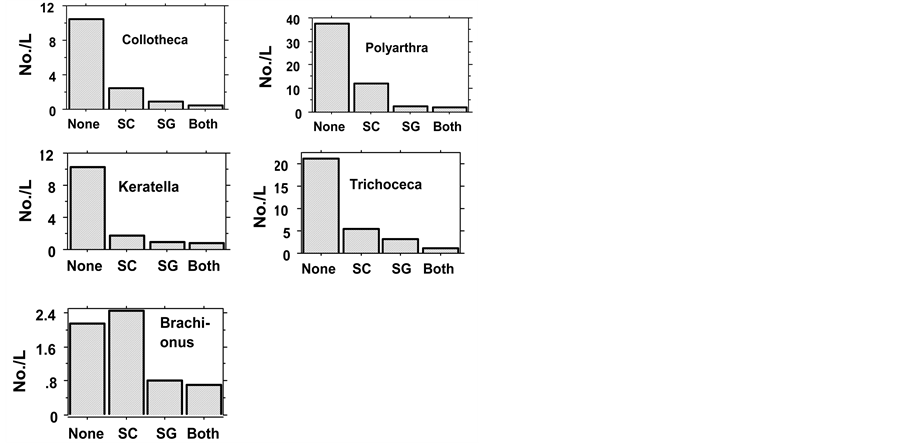
Figure 4. Mean (two experiments all samples) of Rotifera in Fishless (None), H. molitrix (SC), S. galilaeus (SG), SC + Sc (Both). Probability values are given in Table4

Table 1. Comparispns (Kolmogorov-Smirnov Two Samples Test) between treatments SG, SC, Both, None) means, of Initial Day and 12 day. Probability (p) values are presented: SG = S. galilaeus; SC = H. molitrix; Both = Two species together; None = Fishless; NS = Not Significant; S = Significant.
experiments (Figure 1) shows a decrease in day 3 of total phytoplankton chlorophyll in tanks containing either fish species. The presence of fish was also associated with subsequent increase in both net and nano-chlorophyll concentrations at the conclusion of each experiment. Particle size-frequency distribution for experiment 2 (Figure 5)
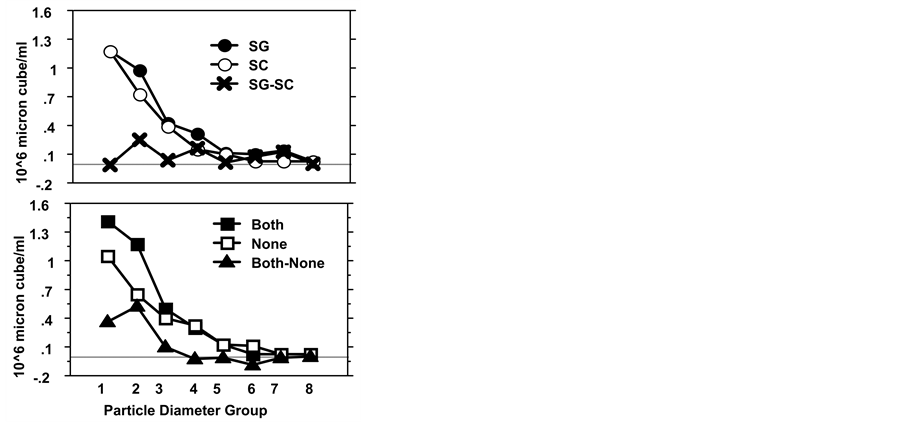
Figure 5. Mean sestonic biomass (106 µ3/ml) estimates values derived from particle counts by Coulter Counter at the end of the second experiment: Upper panel, treatments of S. galilaeus (SG), H. molitrix (SC) and SG minus SC. Lower panel treatments of both species (Both), Fishless (None) and Both minus None. Values presented are the product of counts for each size interval multiplied by the volume of a sphere having a diameter equivalent to that of the mean of the interval. Particle diameter groups are numbered from 1 to 8 when, 1 = 5 − 10 µ, 2 = 10 − 15 µ, 3 = 15 − 20 µ, 4 = 20 − 25 µ, 5 = 25 − 30 µ, 6 = 30 − 35 µ, 7 = 35 − 40 µ, 8 = 40 − 50 µ.
Table 2. Treatment means of population dynamics parameters derived from Ceriodaphnia enclosures for the two experiments (see text): Eg/F = Eggs/Female; Ep/F = Ephipia/Female and r collected on the day of experiment conclusion (12). SG = S. galilaeus; SC = H. molitrix; Both = Two species together; None = Fishless; Exp. 1, 2 = first and second experiment; P = Probability (ANOVA test, S = Significant when p < 0.05) that the parameter value differs from the fishless treatment.
Table 3. Probability values for Sarotherodon galilaeus (SG) and Hypophthalmichthys molitrix (SC) main effects and Interactions (Int.), in two experiments( I, II) for Copepoda and Cladocera.
indicate that significant increase in particle size ranging from 5 to 15 µm EquivalentSpherical-Diameter (ESD) occur in the presence of SG but not with SC. Kolmogorov-Smirnov two sample tests detected significant differences only between those tanks having SG (SG and both) and those not having SG (SC and fishless). SG enhanced
Table 4 . Probability values for Sarotherodon galilaeus (SG) and Hypophthalmichthys molitrix (SC) main effects and Interactions (Int.), in two experiments (I, II) for Rotifers.
the proportion of algal biomass present in the smaller size classes as well as increasing the overall level of production (Figure 5). All treatments also showed significant differences between samples from the beginning and the end of the experiment, while no difference between the particle distribution was detected between treatments on the initial day. Examination of Figure 1 suggests that the effect of either fish on primary production is complex. Highest rates of primary production were associated with the treatments containing both fish species, suggesting a possible correlation with fish biomass (Figure 1). There was an increase of the chlorophyll specific rate of production until day 6 and diminishing later on (Figure 1). SG significantly increased rates of net (p = 0.036 and p = 0.029 for experiment 1 and 2 respectively) and gross primary production (p = 0.063 and p = 0.083 in experiments 1 and 2 respectively). SG also significantly increased chlorophyll specific gross primary production (mg O2 hr−1 microgram chlorophyll−1) in the first experiment (p = 0.036), (Figure 1). Although mean rates of primary production were increased in tanks containing SC there is no significant effect (p = 0.454, and p = 0.509 for experiments 1 and 2 respectively) (Figure 1). Because primary production is a second-order community response, affected by nutrient availability, algal biomass and species composition, it can be expected to exhibit complex interactions with these and other factors, even in short-term experiments.
Ceriodaphnia Enclosures
In the first experiment, Ceriodaphnia enclosures were present only in tanks having SG alone or in fishless tanks. Ceriodaphnia had significantly greater intrinsic rates of increase (r), more eggs per female and fewer ephipia produced in the presence of SG (Table 2). The value of r is derived from the exponential population growth equation:
![]()
where:
N0 and Nt = the populations at the beginning (0) and end (t) of the time interval.
r = Intrinsic rate of increase.
In the second experiment, Ceriodaphnia in enclosures in tanks with SG and with SC had significantly higher values of r (Table 2). Those in tanks with SG also produced fewer ephipia.
4. Discussion
The potential impact of SC on plankton communities, and water quality in general and in particular by suppression of Microcystis biomass and declining of the toxic microcystins was widely studies [5] [9] [12] [13] -[17] and others. Recently a “Chinese carp” invaded and intensively reproduced within the drainage basin of the Mississippi river (USA). The president of USA signed a legislation recommended by the USGS of Chinese Carp ban of stocking. This invader is not the Silver Carp specimen (H. molitrix) (SC) but Bighead Carp (Aristichthys nobilis) (Richardson, 1845) which is well known as zooplanktivorous fish. The ecological destruction concerns with regard to the Bighead Carp in northern USA are justified due to the optimal hydrological conditions for the reproduction of this fish which require long route of river turbulentic flow for the eggs incubation. Nevertheless river Jordan is much shorter with insufficient discharge during most of the time. After 45 years of SC planting history in Lake Kinneret there is not any sign or documentation of reproduction of this fish in the lake. Spataru and Gophen [18] documented more than 50% content of zooplankton in the diet of SC only during 4 summer months. Moreover, Shapiro [19] published data about SC food content in Lake Kinneret as totally (>90%) dominated by phytoplankton. In [5] a data is given about selective feeding of SC on Microcystis (93%) and lower level (7%) of zooplanktivory in the East Lake in China. Miura [5] documented dominance of small size phytoplankters and zooplankton enhancement in East Lake waters as a result of intensive consumption of previously dominant Microcystis by SC. Based on metabolic analysis Miura [5] concluded that the nutritional value of Microcystis is better than that of chlorophytes. Use of confined SC can be recommended to reduce Chl a concentrations in bodies of water where a large percentage of the phytoplankton is in the net-plankton fraction (Laws and Wesburd unpubl. Data). Analysis of Δ13C was done on fishes and plankton collected during winter-spring period and the results were (o/00), (±SD) [20] ): Peridinium-20.6 (1.7); small algae excluding Peridinium—28.8 (2.0); zooplankton—26.2 (4.2); Bleak fishes—25.3 (1.6); adult SG—22.8 (1.2); adult SC—28.0; Detritus—27.3 (0.9). Three consequent conclusions probably emerge from these data: 1) major food component of SC is phytoplankton excluding Peridinium and detritus and secondary status is given to zooplankton; 2) Major food component of Bleaks is zooplankton; 3) Major food component of SG is Peridinium. Filter-feeding fish have complex effects on plankton communities. They may directly suppress zooplankton and phytoplankton prey, while simultaneously indirect influencing taxa which may interact with the primary prey (Table 1, Table 3, Table 4; Figures 2-4). Previous studies have shown that SG suppress zooplankton and probably phytoplankton biomass as well [21] -[23] . Similarly, SC also suppresses zooplankton and has variable effects on phytoplankton, including suppression, no effect, or biomass enhancement [17] [24] -[31] . A prominent direct effect observed over the 12-day course of the two experiments was the elimination of zooplankton by both fish species (Table 1, Table 3, Table 4; Figures 2-4). Impacts of the fishes were not identical however, and final zooplankton densities were higher in treatments containing SC (Table 3, Table 4). This effect was most apparent with rotifers, and may have resulted from several processes, including reduced predation of these small organisms by SC and/or enhancement of rotifers production. Direct feeding effects also probably account for the rapid decline of total chlorophyll concentration in the presence of either fish species during the first three days of each experiment. However, there was an increase in nano-plankton chlorophyll concentration in the presence of either fish during the second half of the experiment. It is suggested that these data might be the evidence of both direct and indirect effects of fish grazing. The decline of total chlorophyll during the first 3 days occurred as the fishes consumed net-phytoplankton together with larger nano-plankton. A secondary effect of the fish feeding is the re-mineralization of nutrients, particularly inorganic nitrogen compounds [22] . The combination of reduced zooplankton grazer densities (i.e. decline of grazing pressure) and increased nutrient availability may have promoted enhancement of nano-plankton abundance during the second half of the experiments. Similar effects were documented, [12] [13] [25] [26] [29] [30] [32] . In fishless tanks the nano-plankton chlorophyll concentration initially increased but then decreased by the conclusion of the experiment. This decline was associated with zooplankton density enhancement and likely resulted from their grazing activities.
Elimination of zooplankton by fish should reduce grazing rates on small-sized phytoplankton. Although there is some overlap in size range of particles ingested by filter-feeding zooplankters and by SG, the particles consumed by zooplankton generally are smaller than 20 µm while those consumed by fish are usually larger than 10 - 20 µm [21] . Particle size-frequency distributions show an increase of concentrations of particles ranging from 5 to 15 µm in tanks containing both fish species. However, the impacts of SG were of greater magnitude than those of SC (Figure 5, Table 1). Test-t (paired) analysis was done for particle size distribution of all treatments and no significant differences were indicated. Nevertheless, ANOVA (p < 0.05) comparison test was done for two particle size groups: <20 µm and >20 µm and the 4 treatments. In all treatments sizes of <20 µm the biomass was significantly higher than that of >20 µm (p values varied between 0.0100 - 0.0062). Consequently it is suggested that beside natural decline of large particles there was an additive effect by fish. Although both fish species reduce zooplankton populations, their effects are not identical, and the responses of specific phytoplankton population to the presence may differ significantly. The large number of significant interaction terms detected in the data (Table 1, Table 3, Table 4, Figures 2-4) suggest that the fish are not simply additive in their impacts on the Lake Kinneret plankton community, and that the impacts of adding SC to the lake community are difficult to predict. Their effect will not be equivalent to the addition of an equal biomass of SG. Although the two fish species are not ecological equivalents, there is still a high degree of similarity in their direct impacts on zooplankton populations. It is likely that SG and SC compete for limited zooplankton resources and stocking of SC may displace SG from some zooplankton resources. The higher intrinsic rate of increase (r), greater number of eggs per female, and lower numbers of ephippia produced in tanks with fish indicate improved conditions for Ceriodaphnia, or in general, filter-feeding zooplankton, relative to tanks without fish (Table 2). These data suggest that the Ceriodaphnia in the experimental enclosures were able to utilize the enhanced nano-phytoplankton populations in the tanks containing fish, and that fish presence actively stimulates secondary productivity among any surviving zooplankton grazers. It can be conclusively summarized that the data presented here indicates that both fish species are direct consumer of zooplkankton and net-phytoplankton. The indirect stimulation of primary production shifts phytoplankton species composition toward a more nano-phytoplankton-dominated community (see also [5] ). Nano-phytoplankton is stimulated because they are not efficiently grazed by the fishes, and because they also experience reduced predation rates from the diminished zooplankton populations. Ceriodaphnia enclosures indicate that any surviving filter-feeding zooplankton should experience increased productivity due to the increased abundance and productivity on nano-phytoplankton food resources. Grazing by the fishes may also rapidly recycled nutrients and contributed to increase in phytoplankton production.
If fish are consuming zooplankton and net phytoplankton while stimulating nano-phytoplankton their impacts on zooplankton populations should be dependent upon fish density. At low densities it might be possible to enhance zooplankton production without major shift to a nano-phytoplankton dominated algal community. In high fish density the dominant effects might include stimulation of nano-phytoplankton productivity and altered specie composition.
5. Conclusion
Lake Kinneret ecosystem has undergone ecological changes. From late 1960’s to early 1990’s, the Kinneret was a phosphorus limited ecosystem and Peridinium was dominant in winter-Spring season with nano-phytoplankton dominance during Summer-Fall months. Stocking of SC in the lake in such conditions is not recommended. The results and conclusions of the present study justify it. Nevertheless, from late 1990’s, the Kinneret ecosystem is Nitrogen limited, Peridinium disappeared and Cyanophyta was enhanced particularly Microcystis. In such conditions, the SC might have the benefit of Microcystis removal. Therefore, during 1970-1990, stocking of SC was correctly objected whilst later on its stocking is justified. During 1970-1990’s enhancement of SG, reducing pressure from zooplankton predation and consequently no SC stocking were therefore recommended. Later on reduction of Microcystis biomass, Micricystins concentration is recommended to improve water quality. The stocking of SC is beneficial for the income of fishermen in both periods.
Acknowledgements
This research was supported by National Science Foundation Research Grant No. BSR-8416519. The first draft was prepared by Dr. Gary L. Vinyard as the first author in cooperation with Dr. Ray W. Drenner and the author, very sadly, G. L. Vinyard deceased. Updating and completing of the MS were done by the author. The paper is dedicated to the memory of G. L. Vinyard, an outstanding scientist, friend, and collaborator.
References
- Gophen, M., Smith, V.H., Nishri, A. and Threlkeld, S.T. (1999) Nitrogen Deficiency, Phosphorus Sufficiency, and the Invasion of Lake Kinneret, Israel, by N2-Fixing Cyanobacterium Aphanizomenon ovalisporum. Aquatic Sciences, 1, 1-14.
- Gophen, M. (2004) Ecohydrological Management of Lake Kinneret: A Case Study. Ecohydrology and Hydrobiology, 4, 397-408.
- Gophen, M. (2004) Hydrology and Management of Lake Kinneret Aimed at Water Quality Protection. In: Zreiny, F. and Jaeschke, W., Eds., Water in the Middle East and in North Africa: Resources, Protection, and Management, Springer-Verlag, Berlin, 207-226. http://dx.doi.org/10.1007/978-3-662-10866-6_18
- Gophen, M. (2008) Long term (1970-2001) Eco-Hydrological Processes in Lake Kinneret and its Watershed. In: Zereini, H., Ed., Climatic Changes and Water Resources in the Middle East and in North Africa, Invited Chapter, Springer, Berlin, 373-402.
- Miura, T. (1989) East Lake: A Phytoplanktivorous Fishes Dominated Lake Ecosystem. Kyoto University. Otsu Hydrobiological Station, 142 p.
- Cindy, S.K., Chapman, D.C., Courtenay, W.R., Housel, Jr., C.M., Williams, J.D. and Jennings, D.P. (2005) Asian Carps of the Genus Hypophthalmichthys (Pisces, Cyprinidae): A Biological Synopsis and Environmental Risk Assessment. Report to US Fish and Wildlife Service per Interagency Agreement 94400-3-0128.
- Gophen, M. (1985) The Management of Lake Kinneret and Its Drainage Basin. Scientific Basis for Water Resources Management, Proc. Jerusalem Symp. IAHS Publ. 153, 127-138.
- Gophen, M. (1987) Fisheries Management, Water Quality and Economic Impacts: A Case Study of Lake Kinneret. In: Shcmidtke, N.W., Ed., Toxic Contamination in Large Lakes. Vol. II: Impact of Toxic Contaminants on Fisheries Management, Lewis Publishers, Boca Raton, 5-24.
- Zhang, X., Xie, P., Hao, L., Guo, N., Gong, Y., Hu, X., Chen, J. and Liang, G. (2006) Effects of Planktivorous Silver Carp (Hypopohthalmichthis molitrix) on Plankton and the Hepatotoxin Microcistins in an Enclosure Experimentin a Eutrophic lake, Lake Shichalai in Beijing. Aquacultur, 257, 173-186.
- Strickland, J.D.H. and Paesons, T.R. (1972) A Practical Hand Book of Seawater Analysis. 2nd Edition, Bull. Fish.Res. Board Can., 167, 310 p.
- Wilkinson, L. (1984) Systat Statistical Package. Systat Inc., Evanston.
- Domaizon, I. and Devaux, J. (1999) Experimental Study of the Impact of Silver Carp on Plankton Communitie of Eutrophic Villerest reservoir (France). Aquatic Ecology, 33, 193-204. http://dx.doi.org/10.1023/A:1009984713183
- Domaizon, I. and Devaux, J. (1999) Impact of Moderate Silver Carp Biomass Gradient on Zooplankton Communities in a Eutrophic Reservoir. Consequences for the Use of Silver Carp in Biomanipulation. Comptes Rendus de Academie des Siences, 322, 621-626.
- Kajak, Z. (1977) Feeding Habits of Silver Carp Hypophthalmichthys molitrix (Val.) and the Problem of Clean Water. Wiad. Ekol., 23, 258-268.
- Miura, T. (1990) The Effects of Planktivorous Fishes on the Plankton Community in a Eutrophic Lake. Hydrobiologia, 200-201, 567-579. http://dx.doi.org/10.1007/BF02530374
- Starling, F.L.R.M. (1993) Control of Eutrophication by Silver Carp, (Hypophtalmichthys molitrix) in the Tropical Paranoa Reservoir (Brasilia, Brazil): A Mesocosm Experiment. Hydrobiologia, 257, 143-152. http://dx.doi.org/10.1007/BF00765007
- Starling, F., Beveridge, M., Lazzaro, X. and Baird, D. (1998) Silvercarp Biomass Effects on the Plankton Community in Paranoa resetrvoir (Brazil) and an Assessment of Its Potential for Improving Water Quality in Lacustrine Envinronments. International Review of Hydrobiology, 83, 499-507.
- Smith, W.D. (1989) The Feeding Behavior of Silver Carp, Hypophtalmichthys molitrix, (Val). Journal of Fish Biology, 34, 817-828. http://dx.doi.org/10.1111/j.1095-8649.1989.tb03366.x
- Spataru, P. and Gophen, M. (1981) Feeding Behavior of Silver Carp Hypophthalmichthys molitrix (Val) and Its Impact on the Food Web in Lake Kinneret, Israel. Hydrobiologia, 120, 53-61. http://dx.doi.org/10.1007/BF00034590
- Shapiro, J. (1985) Food and Intestinal Contents of the Silver Carp, Hypophthalmichthys molitrix (Val) in Lake Kinneret between 1982-1984. Bamidgeh, 37, 3-18.
- Zohary, T., Erez, J., Gophen, M., Berman-Frank, I. and Stiller, M. (1994) Seasonality of Stable Isotopes within Lake Kinneret Food We. Limnology and Oceanography, 39, 1030-1043. http://dx.doi.org/10.4319/lo.1994.39.5.1030
- Drenner, R.W., Hambright, K.D., Vinyard, G.L., Gophen, M. and Pollingher, U. (1987) Experimental Study of SizeSelective Phytoplankton Grazing by Filter-Feeding Cichlid and the Cichlid’s Effects on Plankton Community Structure. Limnology and Oceanography, 32, 1140-1146. http://dx.doi.org/10.4319/lo.1987.32.5.1138
- Vinyard, G.L.R., Drenner, W., Gophem, M., Pollingher, U., Winkelman, D.L. and Hamright, K.D. (1988) An Experimental Study of the Plankton Community Impacts of Two Omnivorous Filter-Feeding Cichlids, Tilapia galilaea and Tilapia aurea. Canadian Journal of Fisheries and Aquatic Sciences, 45, 685-690. http://dx.doi.org/10.1139/f88-082
- Gophen, M.R., Drenner, W. and Vinyard, G.L. (1983) Cichlid Stocking and the Decline of Galilee Saint Peter’s Fish (Sarotherodon galilaeus) in Lake Kinneret, Israel. Canadian Journal of Fisheries and Aquatic Sciences, 40, 983-986. http://dx.doi.org/10.1139/f83-124
- Janusko, M. (1974) The Effect of Three Species of Phytoplanktivorous Fish on Alga Development. Polskie Archiwum Hydrobiologii, 21, 431-454.
- Janusko, M. (1978) The Influence of Silver Carp (Hypophthalmichthys molitrix Val.) on Eutrophication Environment of Carp Pond. Part 3: Phytoplankton. Rocz. Nauk. Roln. Ser. H Rybacto, 99, 55-79.
- Opuszynski, K. (1979) Silver Carp Hypophthalmichthys molitrix (Val.) in Carp Ponds. Part 1: Fishery Production and Food Relation. Ekol. Pol. 27: 71-92. Part II: Influence on ecosystem. Ekologia Polska, 27, 117-133
- Burke, J.S., Bayne, D.R. and Rea, H. (1986) Impact of SILVER and Bighead Carps on Plankton Communities of Channel Catfish Ponds. Aquaculture, 55, 59-68. http://dx.doi.org/10.1016/0044-8486(86)90056-6
- Opuszynski, K. (1981) Comparison of the Usefulness of the Silver Carp and the Bighead Carp as Additional Fish in Carp Ponds. Aquqaculture, 25, 223-233. http://dx.doi.org/10.1016/0044-8486(81)90184-8
- Grigierek, E. (1973) The Influence of Silver Carp (Hypophthalmichthys molitrix Val.) on Eutrophication of the Environment of Carp Ponds. Part: 4. Zooplankton. Roc.Nauk. Rolniczych, Seria H.T., 99, 81-90.
- Kajak, Z., Rybak, J.I., Spodniewska, I. and Godlewska-Lipowa, W.A. (1975) Influence of the Planktivorous Fish Hypophthalmichthys molitrix (Val.) on the Plankto and Benthos of Eutrophic Lake. Polskie Archiwum Hydrobiologii, 22, 301-310.
- Kajak, Z., Spondniewska, I. and Wianiewski, F.J. (1977) Studies on food Selectivity of Silver Carp, Hypophthalmichthis molitrix (Val.). Ekologia Polska, 25, 227-239.