1. INTRODUCTION
Pulmonary hypertension syndrome (PHS) is caused by sustained vasoconstriction of the pulmonary artery system during chronic exposure to hypobaric hypoxia at high altitudes [1-3] or at low temperatures [4-6]. Hypoxia produces morphological and physiological changes as well as changes in the expression levels of genes involved in the production of molecules that are important for adaptation [7-9]. In previous research, low levels of endothelial nitric oxide synthase (eNOS or NOS3) mRNA [10,11] and hypoxia-inducible factor 1 alpha (HIF1α) mRNA [12,13] and elevated levels of endothelin 1 (EDN1) mRNA [14-16] have been found in broiler chickens with PHS. The EDN1, eNOS and HIF1α avian genes regulate blood pressure and maintain oxygen homeostasis at the cellular level in a similar manner to the homologous human genes [17].
In Gallus gallus, the EDN1 gene is located on chromosome two (GGA2) (Gene ID: 420854), and the HIF1α gene is located on GGA5 (Gene ID: 374177). The chromosomal location of the eNOS gene is unknown, although its sequence is found within the final assembly of the genome (Gene ID: 776984).
The aim of this study was to identify functional SNPs in the EDN1, HIF-1α and NOS3 candidate genes and to establish a possible association of these SNPs with the susceptibility of broilers to PHS when they are at high altitudes.
2. MATERIALS AND METHODS
2.1. Population Studied
We used Cobb male broilers were obtained from a commercial hatchery. To identify the birds that were susceptible to the syndrome, the broilers were housed from day 1 to day 49 in an experimental shed in the Poultry Building at the National University of Colombia, which is located at an approximate height of 2638 meters above sea level (m.a.s.l.) (barometric pressure = 560 mmHg) in Bogota, Colombia. The housing, feeding, and management conditions that are suggested in the manual of procedures for the commercial line were used. The temperature was kept between 20˚C and 18˚C using a programmable and automatic system. The following vaccination health plan, which was established by the Instituto Colombiano Agropecuario-ICA, was implemented: Gumboro on day 4, New Castle Disease (strain B1) and infectious bronchitis (Massachusetts strain) on day 8, and revaccination against Newcastle disease on day 18.
2.1.1. Sacrifice and Sampling
The slaughter of the chickens was performed in the chicken slaughtering line of the processing plant at the Institute of Food Science and Technology (IFST) at the National University of Colombia (prior approval for the procedure was obtained from the bioethics committee at the School of Veterinary Medicine and Animal Science at the National University, Bogotá). Prior to sacrifice, animals were individually marked on the leg for identification. Additionally, an inspection of the air sacs and lungs was conducted, and animals with any visible injury were excluded. Subsequently, the heart was removed using surgical scissors and forceps and placed in a flask with aqueous 10% formaldehyde at pH 7.4. Using the same method, fresh muscle samples were placed in Eppendorf tubes that were labeled with the tissue type, date, and animal number and stored at −70˚C for genomic DNA extraction.
2.1.2. Determination of Cardiac Index (CI)
The CI is an indicator of pulmonary hypertension in broilers [18] and is calculated as the weight of the right ventricle (RVW) divided by the total ventricular weight (TVW) multiplied by 100 (RVW/TVW × 100). Animals with a CI of greater than 25 were considered hypertensive, and CI < 22 were considered non-hypertensive [19].
2.2. Genomic DNA Extraction
DNA was extracted from frozen tissue using the saltextraction protocol [20]. The concentration and purity of the DNA was evaluated using a fluorometer, iQuib©, according to the manufacturer’s instructions (Invitrogen™, USA). Visualization was performed using 0.8% agarose gels stained with SYRBgreen® (Invitrogen™, USA). The final working concentration was 10 ng/ul of DNA in 0.1X TE buffer.
2.2.1. Primers Design
The primers sets for EDN1 (Gene ID: 420854), NOS3 (Gene ID: 776984) and HIF-1α (Gene ID: 374177) were designed using the sequences from GenBank and the free software PRIMER3 [21]. The primers were synthesized by an external service, Integrated DNA Technologies (IDT) (Canada).
2.2.2. Amplification
The gene segments were amplified using conventional PCR in a total volume of 12.5 µl. The reactions were performed using 2X GoTaq®Green Master Mix (Promega, USA), 8 pmol of each forward and reverse primer, 1.7 µl of milli-Q water, and 50 ng of DNA. Amplifications were performed using an initial cycle of 95˚C for 3 minutes followed by 30 cycles at 95˚C for 30 seconds, 54˚C at 60˚C for 60 seconds, 72˚C at 90 seconds and a final step the one cycle to 72˚C for 10 min. PCR products were visualized on a 1.5% agarose gel stained with 3 µl of SYRBgreen® (Invitrogen™, USA), and the genotypes were subsequently evaluated using polyacrylamide gels (Table 1).
2.2.3. Polymorphism Detection
To identify polymorphism in the amplified fragments, the single strand conformation polymorphism (SSCP) technique was used [22,23] (Table 2).
Development of Gels:
The silver nitrate protocol was used to develop the gels [23]. Genotypes were read, assigned Arabic numerals, and recorded in a database after being confirmed by two readers in addition to the initial reader.
2.3. Genetic Association
To test the hypothesis of an association between genotypes and PHS and to estimate the penetrance, we used a case-control design based on unrelated individuals [24- 26]. The data were analyzed with the free online statistical program Lamp-0.0.9 [27].
2.4. Sequencing
Sequencing of the candidate genes was only performed for pulmonary hypertensive individuals using background expression [15,13] and changes observed in the SSCP gels [22]. These products were sent to an external sequencing service, and the analysis was per-

Table 1. Primer sequences, sizes of the amplified products, and annealing temperatures.

Table 2. Non-denaturing gel conditions for the SSCP technique.
formed using the program phred, version 0.020425 [28]. The NOS3 (Gene ID: 776984), HIF-1α (Gene ID: 374177) and EDN1 (Gene ID: 420854) gene sequences were used as reference sequences.
3. RESULTS
3.1. Cardiac Index
The CIs ranged from a minimum of 11.28 to a maximum of 54. The data were used as selection criteria to determine which birds were non-hypertensive (controls) and which were hypertensive (cases) (Table 3).
The CI and the genotypes were used to sort the database and perform the association study using the program Lamp-0.09 [27].
3.2. Genetic Association
The hypothesis of association between genotypes of the NOS3 (Table 4) and HIF1α genes with the PHS is accepted (lod > 6, p < 0.001), and it is rejected for the polymorphisms found in EDN1 (lod < 6, p > 0.001). For the genes associated, the proportion of hypertensive chickens was higher for the chickens with a heterozygous genotype 1 2 (penetrance: 80% and 85%). The results of the association and penetrance analysis are shown in Tables 4 and 5.
3.3. Sequence Analysis
3.3.1. EDN1
Three SNPs identified in the 150 bp fragment of intron 3 that was amplified from the sequenced animals (Table 6).
An increased frequency of allele 1 was observed, which was characterized by a C to T (C/T) change in the hypertensive population. But none of these changes were significant associated the PHS (Table 4).

Table 3. Frequency of non-hypertensive and hypertensive birds according to the CI.
3.3.2. NOS3
In the 286 bp fragment that was amplified from the first intron of the NOS3 gene, a consensus sequence corresponding to hypoxic response elements (HRE) 5’[ACGTG]3’ and a 5’CACCC3’ box were identified. The polymorphisms identified were within the consensus 5’[ACGTG]3’ sequence. Allele 1, which is characterized by a C to A or T change, occurs more frequently in hypertensive chickens, while a change of a T to A (T/A) or allele 3, is present in a smaller proportion of the nonhypertensive population (Table 7).
3.3.3. HIF1α
Two SNPs were identified in the amplified sequence from intron 6 of the HIF-1α gene. The frequency of the T to C allele in the study population was higher (46%) in the hypertensive broilers (Table 8).
4. DISCUSSION
In previous studies, the low levels of NOS3 [11] and HIF1α mRNA [13] were found in the lungs of broiler exposed to hypobaric hypoxia under natural conditions (2638 m.a.s.l.), and these low mRNA levels were associated with broilers susceptibility to PHS. In the present study we identified polymorphisms significantly associated with susceptibility of broilers (p < 0.001), and these polymorphisms are positioned within sequences regulation transcription levels the NOS3 and HIF1α genes.
We recognized a consensus sequence corresponding to HRE (ACGTG) within first intron NOS3 gene, and polymorphisms are located in HRE. The allele 1 is characterized by C/A or T change and it was the most prevalent allele in the pulmonary hypertensive population (41%) (Table 7). When allele 1 was combined with allele 2 (G/T or A) probability of disease increased 80% in the studied population (Table 4). This increased probability may occur because HRE acts like a cis-regulatory element binding for transcription factors such as HIF1α
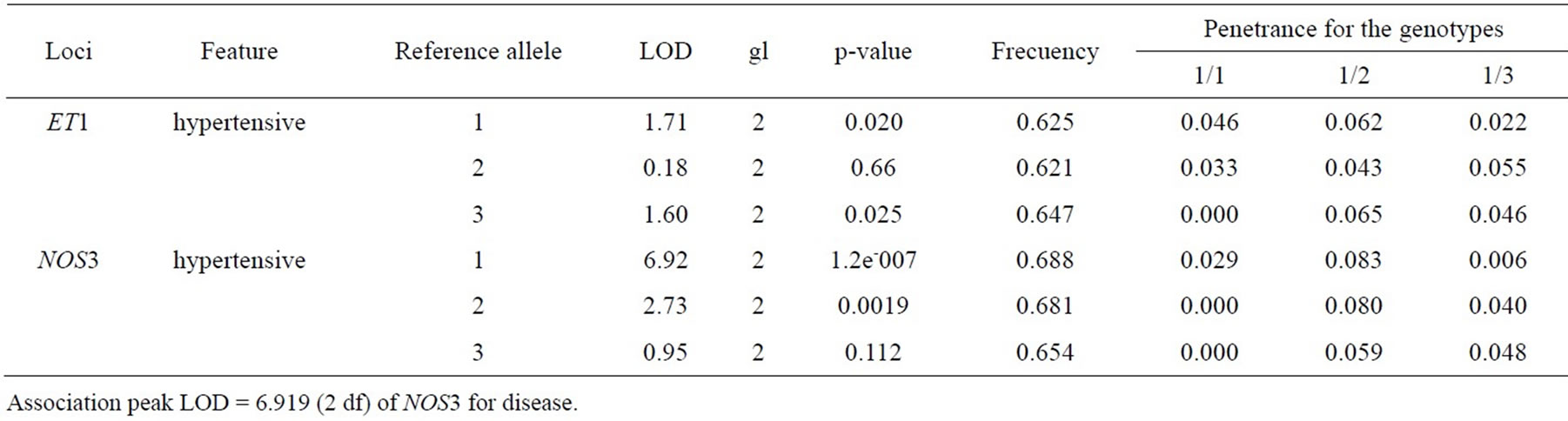
Table 4. Statistical test for the association between the EDN1 and NOS3 genes with PHS.

Table 5. Statistical test for the association between the HIF1α gene and PHS.

Table 6. Polymorphisms identified in intron 3 of the EDN1 gene.

Table 7. Polymorphisms identified in intron 1 of the NOS3 gene.

Table 8. Polymorphisms identified in intron 6 of the HIF1α gene.
[29]. This trans-activation allows organisms to adapt to specific situations such as a low concentration partial of oxygen by increasing the transcription of NOS3 that are dependent on environmental stimulus [30]. The high penetrance for genotype 1/2 (Table 4) assumed from HRE sequence is mutated and trans-activation is not possible, by HIF1α does not recognize the DNA-binding site, and it’s not able to induce gene transcription in hypoxia condition.
Consequently the level of eNOS in pulmonary hypertensive chickens decreases. These changes could be used as genetic markers to predict the susceptibility of broilers to develop pulmonary hypertension.
Decreased levels of eNOS expression in chickens are correlating with low levels of enzyme activity in the pulmonary vessels [10,11,31]. Many studies have shown that changes in levels of endothelial nitric oxide synthesis are implicated in endothelial dysfunction in humans [32] increased your susceptibility to cardiovascular disease and hypertension [32-34], to stroke and kidney disease [33]. Those SNPs change the susceptibility of broilers to PHS at high altitudes and decrease the efficiency of transcription when the chickens are exposed to chronic hypoxic conditions.
The HIF1α avian transcription factor is a product of alternative splicing (Ensembl: ENSGALG00000011870). One in three of these events introduces premature termination codons (PTC) [35] or activates cryptic alternative splice sites [36] that are associated with disease or susceptibility to disease [35,37]. Allele 1 is the most prevalent allele in the hypertensive population (46%); this T/C change (Table 8) may activate a cryptic alternative splice site or change the sequence cis-regulatory elements that regulate alternative splicing. The sequences are recognized by small nuclear particles known as ribonucleoproteins or small nuclear ribonucleoproteins (snRNPs), and these signals regulate editing alternative splicing [36]. In both cases, these cis-regulation elements are located in the introns control mRNA stability, translation efficiency and localization [35]. Hu et al. (2003) [7], Huang et al. (2004) [38] and Shohet and Garcia (2007) [39] have reported that any alteration in the sequence of the alpha subunit of the HIF1α gene can affect transcriptional factor regulation and cause problems during embryonic development in chickens and mammals. Furthermore, changes in the sequence can have a strong impact on cardiac development. Areiza (2010) [13] reported that the levels of HIF1α mRNA are reduced in the lungs of non-hypertensive and hypertensive broilers that are subjected to natural hypobaric hypoxia. One possible hypothesis that could explain this finding is that nonhypertensive birds adapt easily [9,40,41]. However, broilers with pulmonary hypertensive may be susceptible to disease because the changes found in the 6th intron of HIF1α may lead to rapid degradation of the factor by NMD, as suggested by Garcia-Blanco et al. (2004) [42] and Wang and Cooper (2007) [35].
The lack of an association between PHS and alleles identified in the EDN1 gene does not mean that the gene is not involved in the susceptibility of chickens to develop hypoxic pulmonary hypertension. Rather, it means that examining other regions of the gene for polymorphisms that may be altering trans-activation or cis elements is necessary. Pulmonary hypertension is a disease of complex traits that involves the interaction of genetic, epigenetic and environmental factors. In addition, EDN1 is one of the candidate genes involved in regulation of crucial mechanisms that are related to a predisposition to develop the PHS in broilers and humans. Therefore, EDN1 should not be excluded from future research.
The findings of this research indicate that intronic sequences should be included in the search for functional polymorphisms that produce physiological changes. As it is evident in the present study, introns have transcriptional regulatory sequences or signals from post-transcriptional control [43]. Modification of these sequences known as cisand trans-activation elements, can alter the physiological processes of adaptation to hypoxia, especially broiler chickens susceptible to PHS at high altitudes.
With respect to the control birds, its reference genotypes were not sequenced, because, according to previous work, the non-hypertensive birds maintain normal function and expression levels of the HIF1α and NOS3 genes [10,11,13,31], which is why we decided to sequence only pulmonary hypertensive animals.
Although determining the pattern of inheritance of complex diseases is not completely possible, from these results, it can be concluded that the inheritance pattern of PHS is autosomal overdominant with deleterious effects that are characterized by a higher penetrance in heterozygous broilers than in homozygous (Tables 4 and 5). Furthermore, these SNPs result in a decreased ability of broiler chickens to adapt to hypoxic conditions, phenotype caused by changes in the transcript levels of genes involved in adaptation to chronic natural hypobaric.
Finally, we can say that domestic chickens can be used as a research biomodel to advance genetic studies of pulmonary hypertension in humans because the clinical findings in humans and chickens are similar and the cisand trans-activation regulatory elements of the evaluated candidate genes are conserved.
5. ACKNOWLEDGEMENTS
We thank the Research Division of Bogotá (DIB) at the National University of Colombia, which funded this work, and Dr. Diana Álvarez and Susan Castro for her support and collaboration.