Association between human glutathione S-transferase omega rs4925 polymorphism and bladder cancer ()
1. INTRODUCTION
Bladder cancer is a multi-factorial malignancy influenced by environmental risk factors and molecular variations in metabolism-related genes leading to DNA instability and resulting in the development of cancer [1]. Cigarette smoking is a major risk factor for bladder cancer [2,3]. Cigarettes contain several carcinogens includeing polycyclic aromatic hydrocarbons, aromatic amines and N-nitroso compounds which have been reported to be associated with bladder cancer [4]. Epidemiological studies have indicated that cigarette smokers have a significantly higher bladder cancer risk [3,5]. Our previous study also found that cigarette smokers have a significantly increased risk than those who had never smoked [6]. Other risk factors including arsenic exposure through drinking well water, occupational exposure to carcinogens and chronic inflammation are known risk factors for bladder cancer [7,8].
Carcinogens contained in cigarettes generally required the detoxification by phase II enzymes leading to lower toxic and more hydrophilic derivatives and then more readily excreted. However, the deficiency in detoxification-related enzymes generated oxidized products includeing reactive oxygen species (ROS) which can cause DNA damage and the accumulation of genetic mutations [9]. Several genetic polymorphisms of detoxificationrelated enzymes such as glutathione Stransferases (GSTs) which have been shown to be responsible for catalyzing several chemical carcinogens and may modulate the individual susceptibility to the development of various malignancies [10-12].
GST omega 1 (GSTO1) not only carried a cysteine at the active site but also had a specific N-terminal extension of 11 - 20 amino acid residues [13]. GSTO1 also have thioltransferase and dehydroascorbate reductase activities [14]. In addition, GSTO1 can protect cells against calcium-induced apoptosis [15]. A single nucleotide polymorphism (rs4925) of GSTO1 gene was reported at nucleotide 419 causing an alanine (Ala) to aspartate (Asp) substitution in amino acid 140. This amino acid substitution of GSTO1 may lead to a lower enzyme activity of the variant enzyme in a substrate-specific manner [16]. In addition, the GSTO1 Asp/Asp gene genotype not only caused a substitution in amino acid from hydrophobic to hydrophilic residue but also exhibited the thiol-transferase activity only 75% of that of the Ala/Ala genotype [15-17]. Thus, certain genetic variations located in GSTO1 gene might explain the individual susceptibility to oxidative stress from cigarette smoking.
To explore the potential effect of the rs4925 polymorphism of GSTO1 gene on the risk of bladder cancer, we carried out a hospital-based case-control study to address this issue. Furthermore, we tested the joint effect of cigarette smoking and the rs4925 polymorphism of GSTO1 gene on the development of bladder cancer.
2. MATERIALS AND METHODS
2.1. Study Population
A total of 300 histopathologically confirmed bladder cancer (BC) patients were enrolled from the Department of Urology, National Taiwan University Hospital between September 2002 and May 2009. The diagnosis was performed following the routine urological practices, including endoscopic biopsy or surgical resection of urinary tract tumors, with tissue samples confirmed by board-certified pathologists. In addition, a total of 300 cancer-free controls recruited from a hospital-based pool including those who received a general health examination at Taipei Medical University Hospital and Taipei Municipal Wan Fang Hospital. These controls were matched to BC patients in terms of age ±3 years, as well as gender, and had no history of malignancy. All participants provided informed consents and then were interviewed by a well-trained interviewer using a structured questionnaire to collect information including a history of cigarette smoking. Study subjects who had smoked more than 100 cigarettes during their lifetime were regarded as ever smokers, while those who had smoked less than 100 cigarettes were defined as never smokers. The number of pack-years was calculated using the formula: pack-years = 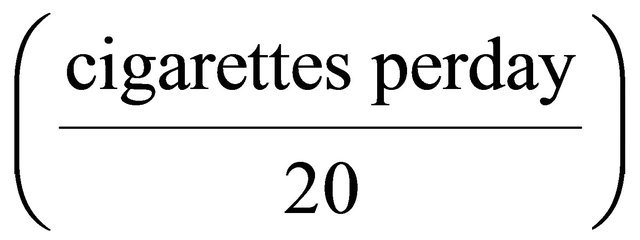 ´ (smoked years). The Research Ethics Committee of the National Taiwan University Hospital, Taipei, Taiwan, approved this study, and this study also complied with the World Medical Association Declaration of Helsinki.
´ (smoked years). The Research Ethics Committee of the National Taiwan University Hospital, Taipei, Taiwan, approved this study, and this study also complied with the World Medical Association Declaration of Helsinki.
2.2. Genotyping of GSTO1 Ala140Asp Polymorphism
Genomic DNA was extracted from the buffy-coat of peripheral blood using standard methods and stored at –80˚C. Genotyping was determined using a polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) method. Briefly, the following primer sequences were designed for the GSTO1 Ala140Asp polymorphism: 5’-GAACTTGATGCACCCTTGGT-3’ (forward) and 5’-TGATAGCTAGGAGAAATAAT TAC-3’ (backward). The PCR conditions were: one cycle at 95˚C for 5 min; 35 cycles of 95˚C for 30 sec, 58˚C for 30 sec and 72˚C for 45 sec, and a final extension at 72˚C for 10 min. The amplified PCR product was 254 bp and visualized by electrophoresis in a 2% agarose gel. After the complete digestion with the restriction enzyme Cac8I at 37˚C for 18 hours, the resulting DNA fragments which represented the Ala140Asp polymorphism of GSTO1 gene were analyzed by electrophoresis in a 3% agarose gels. To ensure quality, a random 10% of the samples were genotyped repeatedly.
2.3. Statistical Analysis
Tests of Hardy-Weinberg equilibrium (HWE) are performed using a χ2 goodness-of-fit test. The joint effects of cigarette smoking and the GSTO1 Ala140Asp polymorphism on BC risk were estimated by odds ratios (ORs) and its 95% confidence intervals (CIs) using a multivariate-adjusted logistic regression. All statistical analyses were performed using the Statistical Analysis Software for Windows, version 9.1 (SAS Institute, Cary, NC). P-values of <0.05 were considered statistically significant.
3. RESULTS
3.1. The Distribution of Basic Characteristics
The distribution of basic characteristics for BC patients and controls is shown in Table 1. Among 300 BC patients, the mean age ± SD (standard deviation) was 65.82 ± 13.74 years. Among 300 controls, the mean age ± SD was 64.31 ± 15.33 years. There were no significant differences in the distribution of age, gender and education level between BC patients and controls. We found a significantly increased BC risk in ever smokers (OR = 2.4; 95% CI = 1.5 - 3.9). The median of packyears among controls who had smoked was 30 packyears. Significantly increased BC risks of 2.1 and 2.4 were also found for study subjects who smoked 1 - 30 pack-years and those who smoked more than 30 packyears, respectively. Moreover, we observed that alcohol drinkers had a significantly higher BC risk of 1.9 (95% CI = 1.3 - 3.4).
Table 1. Demographic characteristics of 300 BC patients and 300 controls.
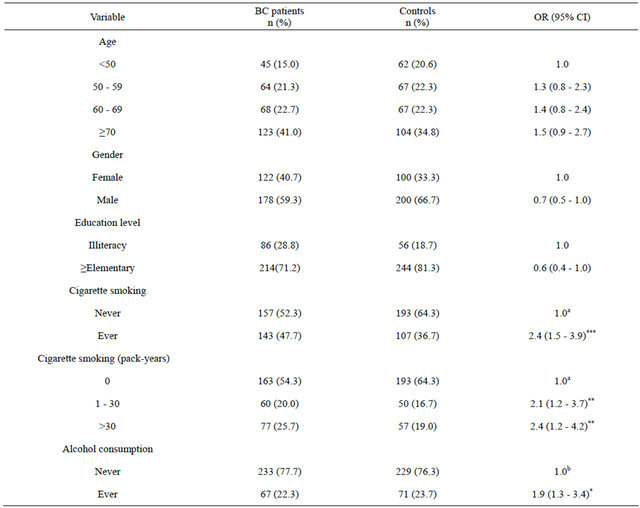
*P < 0.05, **P < 0.01, ***P < 0.001; aAdjusted for age, gender, education level and alcohol consumpti; bAdjusted for age, gender, education leveland cigarette smoking.
3.2. Comparison of the GSTO1 Ala140Asp Polymorphism
The distribution of the observed genotype frequencies among the control group was in HWE for the GSTO1 Ala140Asp polymorphism (P = 0.528). Study subjects who carried the Asp/Asp genotype of GSTO1 gene had a non-significant higher BC risk (OR = 1.1; 95% CI = 0.4 - 4.6) comparing to individuals who carried the Ala/Ala genotype. However, compared with study subjects who carried the combination of Asp/Asp and Ala/Asp genotypes of GSTO1 gene, those with the Ala/Ala genotype had a significantly higher BC risk of 1.5 (95% CI = 1.1 - 2.7) (Table 2).
3.3. Combined Effects of Smoking and the GSTO1 Ala140Asp Polymorphism
Because we hypothesized that single nucleotide polymorphisms located in GSTO1 gene would modulate the toxic effect of chemical carcinogens in cigarettes on BC, we further examined the combined effect of the GSTO1 Ala140Asp polymorphism and cigarette smoking. Comparing with never smokers who carried Asp/Asp and Ala/Asp genotypes of the GSTO1 gene as the reference group, significantly increased BC risks of 1.9 (95% CI = 1.1 - 3.1), 3.1 (95% CI = 1.5 - 6.5) and 4.9 (95% CI = 2.8 - 9.7) were found for never smokers carrying the Ala/Ala genotype, ever smokers with Asp/Asp and Ala/Asp genotypes and ever smokers with the Ala/Ala genotype, respectively (Table 3).
4. DISCUSSION
Chronic exposure to chemical carcinogens from cigarette smoking can lead to DNA damage adducts [9]. In consistent with previous studies, significantly increased BC
Table 2. Distribution of GSTO1 Ala140Asp polymorphism in 300 BC patients and 300 controls.

**P < 0.01; aAdjusted for age, gender, education level, cigarette smoking and alcohol consumption.
Table 3. Combined effect of GSTO1 Ala140Asp polymorphism and cigarette smoking on BC risk.
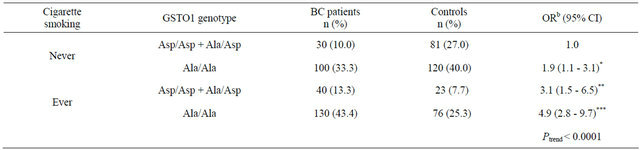
*P < 0.05; **P < 0.01; ***P < 0.001; bAdjusted for age, gender, education level and alcohol consumption.
risks of 2.4, 2.1 and 2.4 were found for ever smokers, light smokers (1 - 30 pack-years) and heavy smokers (>30 pack-years), respectively. In addition, we observed a significantly increased UC risk of 1.9 for alcohol drinkers. Some studies reported that cigarette smoking and alcohol consumption have synergistic interaction on tumorigenesis [18].
GSTO1 has been found in various human tissues [15]. We observed that study subjects who carried the Asp/Asp genotype of the GSTO1 Ala140Asp polymorphism had a slightly higher BC risk. A previous study reported that significantly increased risks of hepatocellular carcinoma, cholangiocarcinoma and breast cancer were found for those carrying the Asp allele of the GSTO1 Ala140Asp polymorphism [19]. A study reported that the enzyme activity of GSTO1 Ala/Asp genotype was 75% of that of GSTO1 Ala/Ala genotype indicating this genetic variation may lead to defective protection against oxidative stresses [15]. However, several studies reported that the GSTO1 Ala140Asp polymorphism would not influence the thioltransferase activity and further modify the risk of malignancy [20-23]. The discrepancies between our findings and previous studies may be due to ethnic variation. The Asp allele frequency was significantly different among various populations. The Asp allele frequency was 0.34 for European population, 0.08 for Africans, 0.17 for Han Chinese and 0.12 for Japanese [19]. Our findings showed an Asp allele frequency of 0.18 which was similar to Asian population such as Han Chinese and Japanese.
The present study also found that a significant joint effect of cigarette smoking and the Ala/Ala genotype of the GSTO1 Ala140Asp polymorphism on BC risk (OR = 4.9), showing a dose-response relationship (Ptrend < 0.0001). Therefore, our results implied that several environmental risk factors should be diminished to prevent individuals from the development of BC. This study has some limitations. Other factors including secondhand smoke exposure may modify the risk of BC; however, lack of detailed information on exposure of environmental tobacco smoke in this study. Moreover, we select only one functional SNP of GSTO1 gene which might not exhibit entire expression and enzyme activity of GSTO1 [24,25]. Therefore, more functional SNPs of GSTO1 gene and other candidate genes should be included in a future study with a larger sample size.
In conclusion, we carried out a hospital-based casecontrol study to investigate the joint effect of cigarette smoking and the GSTO1 Ala140Asp polymorphism (rs4925) on the development of bladder cancer. Our findings showed that a significantly highest risk of bladder cancer was found in subjects who had smoked and carried the Ala/Ala genotype of GSTO1 gene. Therefore, we suggest that the decreasing of environmental risk factors could prevent individuals from the risk of bladder cancer, especially for those who carried the risk genotypes of detoxification-related genes.
[26] NOTES
[28] *These authors contributed equally to this work.
[29] #Corresponding authors.
[30]