Genetic Diversity and Population Structure of North China Mountain Walnut Revealed by ISSR ()
1. Introduction
Several walnut (Juglans spp.) species are important dual purpose fruit and timber economic forest tree species, of these the globally most important is “Common Walnut” (J. regia). China is the one of origin centre of wal- nuts and possesses abundant resources. There are five main species of the Juglans genus available as walnut breeding resources in China. These are J. regia L., J. Sigillata Dode, J. mandshuria Maxim, J. openensis Hu, and J. nigra L. [1] . Due to walnut being seed propagated with open-pollination, walnut is highly variable with a diverse gene pool [2] . Walnuts in China have been divided into four ecotypes (Sinkiang Walnut, North China Mountain Walnut, Qinling-Daba Mountain Walnut and the Tibetan Highlands Walnut) in terms of their geo- graphical distribution [3] . Genetic diversity research on North China Mountain Walnuts has developed slowly in comparison with the other three ecotypes in China.
In China, North China Mountain Walnuts are mainly distributed in the foothills of the North China Mountains, which spread across the northern latitudes from 35˚ to 40˚, and the western longitudes from 112˚ to 119˚. The area is a rocky, mountainous region with serious soil erosion, a warm temperate climate, an annual precipitation from 400 - 700 mm, and slightly calcareous cinnamon soils [1] . Owing to the unique geography and climate, the endemic walnut varieties contain many agronomically excellent properties and specific valuable genes, such as for high content of protein, and strong disease and drought resistances [3] , which have significant potential value for walnut variety improvement. In addition, the North China Mountains could be where J. regia L. and J. mandshurica Maxim were first domesticated and the region is one of the cultivated species’ provenance [3] . Thus, the North China Mountain Walnut could possess a vast gene resource. However, in the past decade, under long-term biological or environmental pressure, the native walnut resources have been seriously damaged, which may result in increased homogeneity or reduction of genetic variability. The lack of systematic studies of genetic diversity among Chinese Juglans species and ecotypes could seriously restrict genetic improvement by limiting exploitation in walnut culture and breeding of many excellent traits found in these landrace lines. Ac- cordingly, it is essential to properly characterize and assess the genetic diversity of landrace walnut resources for protection and breeding utilization.
In the past few years, a quantity of molecular markers, such as RFLP (Restriction Fragment-Length Poly- morphism) [4] [5] , RAPD (Randomly Amplified Polymorphic DNA) [6] -[9] , AFLP (Amplified Fragment Length Polymorphism) [10] [11] , SSR (Simple Sequence Repeat or microsatellites) [9] [12] -[17] , and ISSR (Intersimple Sequence Repeat) [18] have been generally employed to examine genetic diversity and structure in walnut. While previous reports concentrated on the fingerprinting and genetic variability of walnut germplasm, there are few studies on the population genetic diversity of different ecotypes [16] [19] [20] , especially the North China Mountain Walnut. ISSRs have been widely employed to evaluate genetic diversity of populations of economic forest tree species [21] -[23] .
It is reported for the first time to assess genetic diversity and genetic structure of North China Mountain Wal- nut sampled out of eight significant natural populations in China by ISSR. We evaluate the population-level and species-level genetic diversity of eight North China Mountain Walnut populations throughout their known dis- tribution by ISSR analysis to give some references for conservation measures and germplasm resource devel- opment and utilization.
2. Materials and Methods
2.1. Plant Material and Sampling Strategy
In this study, a total of 138 individuals of North China Mountain Walnut, representing eight populations (GX, FY, LY, YX, LC, SX, ZQ, and LZ), were sampled across three provinces in China, including Shanxi, Hebei and Henan along the Taihang and Lvliang Mountains (Figure 1 and Table 1). The collection locations represented a wide geographic distribution of North China Mountain Walnut. Interpopulation distance ranged from a mini- mum of 30.8 km (LC-SX) to a maximum of 512 km (LZ-LY). The number of samples analyzed depended on the usability of adult plants (>80 cm at breast height diameter) in each sampling location. In each population, 4 - 25 adult walnut trees were randomly sampled at the distance exceeding at least 50 m from each other to reduce the probability of sampling from family clusters. From each individual tree, fresh mature leaves were randomly sampled, stored in plastic bags and taken to the lab for experimental studies.
![]()
Figure 1. Geographic locations of sampling sites for the eight natural populations of North China Mountain Walnut ex- amined in this study. The abbreviations of GX, FY, LY, YX, LC, SX, ZQ and LZ follow those used in Table 1.
![]()
Table 1. Location and sampling size of eight nature populations of North China Mountains walnut.
2.2. DNA Extraction
Genomic DNA was extracted from storage leaves of walnut by using the optimized Cetyl Trimethyl Ammonium Bromide (CTAB) method [11] . Total DNA quality was checked by gel electrophoresis and DNA concentration was determined by Biophotometer protein nucleotide analyzer (Eppendorf, Germany). Purified DNA was di- luted to approximately 30 ng∙μL−1 prior to polymerase chain reaction (PCR) amplification, and preserved in the refrigerator at −20˚C.
2.3. ISSR-PCR Amplification
A total of one hundred ISSR primers were used. The primers were designed according to public sequences provided by the University of British Columbia, Canada and biosynthesised by the Shanghai Sangon Biological Engineering Technology & Services Co. Ltd. (Shanghai, China). They were sifted firstly grounded on one sam- ple from each population. Nine primers, which yielded clear, polymorphic, and reproducible bands, were used for the further genetic diversity analysis of all 138 DNA individuals (Table 2). PCR reactions were performed in 25 μl reaction mixture containing 2.5 μl of 10× buffer, 1 U Taq polymerase, 0.2 μM dNTP (Tiangen Biotech (Beijing) Co., Ltd), 0.2 μM primer and 30 ng of genomic DNA. Amplification was carried out in a thermocycler (Peltier Thermal Cycler PTC-200, MJ research, USA) as follows: initial denaturation of 5 min at 94˚C, followed by 35 cycles of denaturation of 30 s at 94˚C, annealing of 45 s at 51˚C - 60˚C (based on the used primer), and extension of
1.5 m
at 72˚C, followed by a final extension of
7 m
at 72˚C. The amplification products were sepa- rated on 2.0% agarose gels in 0.5× TBE buffer at a constant voltage of 80 V, for 2 h along with 100 bp DNA ladder marker (100 - 1500 bp) and stained by 0.1% ethidium bromide. The gel images were then visualized and recorded using a Bio-Rad Gel Doc 2000TM system (USA).
2.4. Data Analysis
Polymorphic bands were scored visually as either present “
1”
or absent “
0”
to form a binary matrix. On the supposition of Hardy-Weinberg equilibrium, POPGENE versions 1.31 [24] were employed to compute the fol- lowing basic statistics. To assess genetic diversity at species level and population level, the parameters calcu- lated included; percentages of polymorphic bands (PPB%), mean number of alleles (Na), mean effective number of alleles (Ne), Nei’s gene diversity index (H) and Shannon-diversity index (I). To evaluate gene differentiation between populations, the parameters calculated included Nei’s coefficient of population differentiation (GST) with the formula: 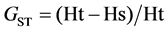 [25] , and Gene flow among populations (Nm) which was calculated us- ing the formula of
[25] , and Gene flow among populations (Nm) which was calculated us- ing the formula of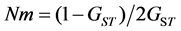 . Genetic relationships among populations were estimated by Nei’s un- biased genetic distances (D) and genetic identities (I) using NTSYSpc-2.0 [26] , and a cluster analysis was per- formed using UPGMA (the Unweighted Pair Group Method with Arithmetic mean) [27] . In addition, population differentiation coefficients within and among the populations were computed from AMOVA 1.55 (analysis of molecular variance) [28] .
. Genetic relationships among populations were estimated by Nei’s un- biased genetic distances (D) and genetic identities (I) using NTSYSpc-2.0 [26] , and a cluster analysis was per- formed using UPGMA (the Unweighted Pair Group Method with Arithmetic mean) [27] . In addition, population differentiation coefficients within and among the populations were computed from AMOVA 1.55 (analysis of molecular variance) [28] .
3. Results
3.1. Genetic Diversity
91 bands in total were generated from nine screened primers for the 138 individual samples from the eight pop- ulations, of which 84 (92.13%) were polymorphic bands. The size of the amplified band ranged from 300 bp to 1200 bp. The percentage of polymorphic band (PPB) of the 9 ISSR primers differed from 66.67% (UBC855) to 100% (UBC815, UBC836, UBC840 and UBC853), with an average of 92.31%. The number of bands presented by each primer also differed from 6 (UBC855) to 11 (UBC840) with an average of 10.11 (Table 2).
Estimates of genetic diversity in eight populations of North China Mountain Walnut are summarized in Table 3.
![]()
Table 2. Polymorphic bands generated by ISSR in eight nature populations of NCMW.
Y = (C,T), R = (A,G), B = (C,G,T), D = (A,G,T). aSequence 5' - 3'; bPCR reaction annealling temperature for tabulated primers (˚C); cPercentage of polymorphic band.
![]()
Table 3. Genetic diversity analysis of 8 populations of North China Mountain based on ISSR markers.
PPB, Percentage of polymorphic loci (%); Na, Observed number of alleles; Ne, Effective number of alleles; H, Nei’s genetic diversity; I, Shannon’s information index; SD, Indicates the standard deviation.
At the species level, the effective number of alleles (Ne), Nei’s genetic diversity (H) and the Shannon informa- tion index (I) were 1.4243, 0.2592, 0.4003 respectively. At the population level, PPB ranged from 19.78% for the LZ population to 57.14% for the YX population, with an average of 43.27%, and the average Ne, H, and I were 1.2253, 0.1347, and 0.1862, respectively. The results showed that the genetic diversity of North China Mountain Walnuts from the YX population (Ne = 1.3163, H = 0.1839, I = 0.2778) was the richest in three esti- mates among the eight populations and the lowest diversity was exhibited in the LZ population.
3.2. Genetic Differentiation
Nei’s coefficient of genetic differentiation (GST) was 0.5066, which showed that 50.66% of the total genetic va- riability is attributed to inter-populations and 49.34% intra-populations. Results of AMOVA also agreed with which genetic differentiation inter-populations is relatively high (FST = 0.5161). At two hierarchical levels, 51.61% of the total genetic variation was partitioned inter-populations, and 48.39% (P < 0.001) intra-populations. The level of gene flow (Nm) was estimated to be only 0.5133, which implied that a low gene flow would not prevent genetic drift thus enabling the gene differentiation between populations.
3.3. Genetic Similary and Genetic Distance among Populations
To further clarify the gene differentiation among different populations, Nei’s pairwise genetic similary coeffi- cients were assessed. Genetic similar coefficients varied from 0.7423 to 0.9298 with an average 0.8159, which showed higher levels of genetic differentiation partitioned among populations of North China Mountain Walnut. The highest similar coefficient was between the ZQ population and the LC population, while difference in simi- lary coefficients between the FY population and the LC population was revealed as the lowest (Table 4).
In order to further illustrated relationships among populations, a dendrogram generated by UPGMA algo- rithm based on Nei’s genetic similary, which clustered the eight populations into three major groups (Figure 2). The first group included five populations (ZQ, LC, SX, YX and LY), distributed along the main vein of Taihang Mountains, the second group comprised GX and FY populations, located in the south of Taiyue Mountains and east of Lvliang Mountains respectively, while LZ alone formed the third group, located in the east of Taihang Mountains. However, the cluster result was not necessarily relationship with geographic separation. Such as, the ZQ population had a much greater geographic separation from the LY population than the LZ population, but ZQ clustered together with the LY population and not with the LZ population in the dendrogram. Therefore, geographical distance alone cannot account for the genetic distance between populations.
4. Discussion
4.1. Genetic Diversity
Our data shows that genetic diversity is higher at the species level (PPB = 92.31%, H = 0.2592, I = 0.4003), but
![]()
Table 4. Nei’s unbiased genetic identity (above diagonal) and genetic distance (below diagonal) among eight nature popula- tions of North China walnut.
![]()
Figure 2. ISSR amplification results with primer UBC811 for GX population.
lower at population level (PPB = 43.27%, H = 0.1347, I = 0.1862), which is alike to the results obtained from same species using different molecular markers [17] [19] [20] . When compared with many other economic tree species using ISSR, North China Mountain Walnut maintains higher species diversity (e.g. gingko [29] , Emme- nopterys henryi [30] , etc.). The higher species diversity may largely be attributed to walnut breeding systems. The walnut is wind pollinated. When compared with insect pollination, the smaller wind carried pollen is easier to spread. In addition, walnut has unisexual flowers and a monoecious plant, which may carry out both geitono- gamy and cross pollination, but due to dichogamy, the same cultivars or strains while self fertile may not be able to pollinate itself, which results in frequent gene flow among individuals and increases the chance of gene re- combination. Geographical distribution would be another key factor that determines genetic diversity level of a species. The North China Mountain Walnut is distributed mostly in rocky Mountainous areas, which have se- rious soil erosion and calcareous cinnamon soils. The need and hence ability to adapt to different environments results in a higher level of genetic diversity.
North China Mountain Walnuts are distributed in different ecological environments and geographic condi- tions, such as having a range of growing elevations from 45 m to 1430 m above sea level. With increasing ele- vation, climate and soil physicochemical properties also show a significant change, which results in greater pop- ulation variation. As observed in our study, the population differentiation parameters such as PPB, Ne, H and I in eight populations showed a similar pattern, the YX population is the highest, and the LZ population is the lowest. This can be interpreted as a consequence of habitat fragmentation and small numbers of individuals. Due to genetic drift, small populations tend to lose genetic variation has been verified in many studies. Therefore, the results need to be further studied and verified by a more extensive investigation.
Compared with other ecotypes, genetic diversity of North China Mountain Walnut was at a moderate level, which was marginally higher than Sinkiang walnut and Qinling-Daba Mountain walnut [31] , but was lower than Tibet walnut [17] . The result of this study supported the conclusion that the Tibet walnut population diversity was generally higher than other ecotypes populations [17] .
4.2. Populations Genetic Structure
In accordance with the GST value (GST = 0.5066), found in this study a significant amount of genetic differentiation is observed among and within eight populations of North China Mountain Walnut. This point is reinforced by the AMOVA, which indicating that 51.61% of the genetic variation was partitioned inter-populations and 49.34% intra-populations. Significant genetic differentiation was also reported by Wu et al. using RAPD. In contrast, less genetic differentiation was found by Xi [3] using a bioecological survey method and peroxidase isoenzymatics marker. The difference might be due to the sampling locations, sampling size, and the different marker systems used. Similar studies have shown that there are inconsistencies in genetic differentiation value found between populations when using different genetic markers in other species populations [32] [33] .
High genetic differentiation of North China Mountain Walnut is attributed to the following points: Firstly, because of low gene flow of the North China Mountain Walnut habitat (Nm = 0.5133), the present population structure was shaped by genetic drift mainly. Secondly, the Taihang and Lvliang Mountains contribute to the geographical isolation, which severely hindered dispersal of pollen or seeds and exchange of genes between populations, ultimately promoting the large genetic differentiation among populations. Thirdly, in order to adapt to the diverse habitats of the region in the evolutionary process, variants to some extent occurred, which may have been preserved and fixed gradually due to limited gene flow, thereby, genetic differentiation among popu- lations occurred. The GST value and AMOVA analysis results obtained in this study indicated that North China Mountain Walnut has a significant genetic differentiation among populations, which suggests that the species has a strong environmental adaptability. Put briefly, high genetic variation among populations is due to genetic drift and geographical isolation of the populations.
Based on the genetic similarity, eight populations of North China Mountain Walnut were grouped into three clusters, but genetic differentiation did not completely cluster according to geographical distances (Figure 3). This supported the conclusion that owing to the complex origin and affiliation of walnut, there isn’t an obvious association between genetic similarity and geographic origin [11] . But the results of cluster analysis did corres- pond with phenetic classification systems of local varieties. Mian walnut, with a less developed endoseptum, a slightly colored and a slightly veiny shell, is the main variety in the LC, ZQ and SX populations; it is characte- rized by large fruit size, thin and smooth shell and pale yellow kernels in the LY and YX population; the Fenz- hou and Guxian walnut is well known to have a thin shell, full kernels, higher kernel crackout, milky white ker- nels in the FY and GX population, respectively; the varieties of the LZ population are Mian walnut and Jia wal- nut with thicker, corrugated and tawny shell and low kernel crackout. Whether these differences are due to ge- netic drift still needs to be determined.
4.3. Conservation Implications
To know about the geographic structure of intraspecific genetic variation is indispensable for the protection of germplasm resources. Our study provides an insight into genetic diversity and genetic differentiation at popula- tion levels of North China Mountain Walnut, and indicates a decrease of genetic diversity caused by genetic drift is unacceptable. Accordingly, so as to secure existing diversity, preservation areas covering large popula-
![]()
Figure 3. UPGMA dendrogram for 8 populations of North China Mountain Walnut based on genetic identity.
tions as well as many small populations should be established. Mixing genetically diverse populations of North China Mountain Walnut in situ or ex situ is unacceptable in conservation.
Acknowledgements
We gratefully thank Professor Chuck Leslie at Department of Plant Sciences, University of California, Davis; and Jihua Tang at Henan Agriculture University, for critically reviewing our manuscript. This work was finan- cially supported by the National Natural Science Foundation of China (31171943), the Key Scientific and Technological Project in Henan province (92101110600), and the Scientific and Technological Project in Henan province (102102110178).
NOTES
*Corresponding author.