 Open Journal of Forestry 2014. Vol.4, No.1, 1-7 Published Online January 2014 in SciRes (http://www.scirp.org/journal/ojf) http://dx.doi.org/10.4236/ojf.2014.41001 OPEN ACCESS Synergic Effect of Mucuna pruriens var. Utilis (Fabaceae) and Pontoscolex corethrurus (Oligochaeta, Glossoscolecidae) on the Growth of Quercus insignis (Fagaceae) Seedlings, a Native Species of the Mexican Cloud Forest María L. Avendaño-Yáñez1*, Ángel I. Ortiz-Ceballos1, Lázaro R. S ánchez-Velásquez1*, María R. Pineda-López1, Jorge A. Meave2 1Instituto de Biotecnología y Ecología Aplicada (INBIOTECA), Universidad Veracruzana, Veracruz, México 2Departamento de Ecología y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Autónoma de México, México D.F., México Email: *luzavend@gmail.com, *lasanchez@uv.mx Received October 19th, 2013; revised November 23rd, 2013; accepted December 9th, 2013 Copyright © 2014 María L. Avendañ o-Yáñez et al. This is an open access article distributed u nder the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In accordance of the Creative Commons Attribution License all Copyrights © 2014 are reserved for SCIRP and the own er of the in tellectual property Marí a L. Av en dañ o-Yáñez et al. All Copyright © 2014 are guarded by law and by SCIRP as a guardian. Propagation of native species in local nurseries is an important activity in reforestation and forest restora- tion programs. A requisite for successful plantation is that nursery produced plants are of a size and qual- ity that allows optimal establishment under field conditions. Manipulation of edaphic processes through the combined use of the earthworm Pontoscolex corethrurus, Mucuna pruriens and inorganic fertilizers may promote faster biomass gain. This study assessed the activity of P. corethrurus, its association with M. pruriens (green manure) and inorganic fertilizers, on the growth of Quercus insignis seedlings under greenhouse conditions. Measured variables were basal diameter, height, biomass and foliar nitrogen con- tent. Growth rates of basal diameter (F = 5.33; P < 0.0001) and height (F = 2.84; P < 0.0087) were sig- nificantly greater in the treatment of P. corethrurus-M. pruriens-inorganic fertilizer, relative to the control. Also, leaf biomass and total biomass of the seedlings were greater in the treatment of P. corethrurus-fer- tilizer (F = 2.32; P < 0.0290, F = 3.71; P < 0.0011, respectively) compared to the control treatment. Foliar nitrogen content was significantly higher (F = 2.54; P < 0.01742) in the treatment of P. corethru- rus-inorganic fertilizer. Incorporating biological soil management techniques in propagation of native species is a good choice to assist reforestation and forest restoration. Keywords: Nursery; Oak Seedlings; Earthworms; Green Manure; Inorganic Fertilizers; Plant Propagation Introduction Cloud forest hosts around 6790 plant species (Villaseñor, 2010). Regrettably, this diversity is in decline due to several factors of disturbance, most of which are anthropogenic (Ramírez-Marcial et al., 2001; Lamb et al., 2005). This situa- tion affects the population dynamics of plant species that are endangered, threatened or susceptible to forest fragmentation (Saunders et al., 1991; Cayuela et al., 2006). This is the case with Quercus insignis, a species native to this ecosystem. De- spite a broad geographic range that spreads from eastern-central Mexico to Costa Rica, this species is a narrow habitat-spe- cialist and is highly susceptible to disturbance (Valencia, 2004). It was included in the Red List of Oaks under the endangered category (Oldfield & Eastwood, 2007). More recently, and due to the accelerated destruction of its habitat, this species was re-classified to the Critically Endangered category in the Red List of Mexican cloud forest trees (González-Espinosa et al., 2011). A potential mechanism to reverse this situation and restore degraded areas is the establishment of forest plantations, or the reintroduction of locally extinct species (Vázquez-Yanes & Cervantes, 1993; Meli, 2003; Pedraza & Williams-Line ra, 2003; Lamb et al., 2005). This process must be complemented by the production of native species in nurseries (Benítez et al., 2002). At present, there is nil to very low availability of native species in local nurseries in Mexico, and the few plants that are pro- duced are lacking in both quality and size (Arriaga et al., 1994; Meza-Sánchez et al., 2009). Moreover, the use of black plastic bags with repeated watering causes the soil compaction and poor root development in the seedlings. Novel propagation techniques are therefore required in order to improve seedling quality in nurseries (Benítez et al., 2002), as well as comple- mentary strategies with which to facilitate the future field es- tablishment in the field. In nurseries that are devoted to the massive production of na- tive forest species, common use is made of organic remains, *Corresponding authors. M. L. AVENDAÑO-YÁÑEZ ET AL. OPEN ACCESS sand, fertilizers and pesticides. However, the use of green ma- nure could become an important source of nutrients (mainly nitrogen) to increase soil fertility (Smyth et al., 1991; Blanchart et al., 2006). The legume Mucuna pruriens var. utilis has been used as a cover crop for these purposes (Hulugalle et al., 1986; Smyth et al., 1991; Buckles, 1995; Ortiz-Ceballos et al., 2012) and its use has also enabled the biological control of weeds, pests and diseases, while it acts to influence the composition and activity of the soil biota, particularly the earthworms. All of these effects may indirectly favor plant growth (Ortiz-Ceballos & Fragoso, 2004; Blanchart et al., 2006). The soil biota is known to regulate the availability of nu- trients necessary for plant growth and development (Wardle et al., 2004). There is also evidence of the important role of earthworms in the functioning of natural- and agro-ecosystems through the conservation of soil fertility (Wardle, 2002; Bha- dauria, 2010) and enhancement of plant growth (Lee, 1985; Haimi & Einbork, 1992; Pashanasi et al., 1992; Edwards & Bohlen, 1996; Doube et al., 1997; Scheu, 2003; Ortiz-Ceballos et al., 2007). However, most information published to date on plant-earthworm interactions is derived from studies based on domesticated plants (Edwards & Bater, 1992; Brown et al., 1999; Scheu, 2003; Ortiz-Ceballos & Fragoso, 2004). It is therefore important to examine the influence of earthworms on plants from natural environments (Scheu, 2003), such as cloud forest. The earthworm Pontoscolex corethrurus (Müller, 1857) is a peregrine species of the Glossoscolecidae family and native to the Neotropics. It is an endogeic earthworm with a broad environmental tolerance that occurs in various habitats and soil types (Lavelle et al., 1987; Lee, 1985; Fragoso et al., 1999; Buch et al., 2011), including cloud forest (Fragoso et al., 1999). This species plays a crucial role in organic matter decomposi- tion as well as the mineralization of nitrogen and phosphorous (Barois et al., 1999; Bhadauria & Saxena, 2010) and has been successfully used in bio-fertilization and bio-stimulation tech- niques, producing significant positive effects on crop yields (Lavelle & Pashanasi, 1989; López-Hernández et al., 1993; Senapati et al., 1999). Obtaining information on the interaction between P. coreth- rurus and native cloud forest plant species in general and Q. insignis in particular may be of great relevance, as this can allow us to develop new practices aimed at supplementing and reinforcing the management and propagation of native plant species within conservation and restoration programs for this mountain ecosystem. The goal of this study was to evaluate integrated soil fertility management, i.e., how the combination of biomass (Mucuna pruriens var. utilis), inorganic fertilizer, and earthworms could impact the soil fertility as well as the growth and leaf nitrogen content of Quercus insignis seedlings under nursery conditions. Methods Study Site and Soil The study was conducted in the greenhouse and laboratory of the Instituto de Biotecnología y Ecología Aplicada (INBI- OTECA) in the center of Veracruz State, a region of Mexico that features cloud forest. A total of 200 kg of soil was col- lected to a depth of 40 cm from the Plan de San Antonio cloud forest (19˚26'N, 96˚59'W; 1430 m elevation), located 20 km from the town of Coatepec city. Soil samples were sealed in plastic bags and transported to the INBIOTECA greenhouse where they were air-dried at ambient temperature. The physical and chemical soil properties of the collected soils were; pH 5.7, organic matter content 16.6%, total nitrogen 0.68%, 0.32 cmol∙kg−1 K, 0.4 cmol∙kg−1 Ca, 1.7 cmol∙kg−1 Mg, 26.9 cmol∙kg−1 cation exchange capacity, 54% moisture content (soil collected), 17% clay, 32% silt, 49% sand. Earthworms, Green Manure and Oak Seedlings Juvenile individuals of P. c orethrurus used in this study cor- responded to the first generation obtained under laboratory conditions from earthworms collected in a secondary cloud forest. Pontoscolex corethrurus is an exotic species that has been reported in Mexican cloud forests and is also commonly found in areas that have been transformed from cloud forest to pastureland or crops, such as maize or beans, among others (Fragoso et al., 1999; Fragoso, 2001). Juvenile P. corethrurus earthworms were raised following the protocol of Ortiz-Ceballos et al. (2005): (1) two adult earthworms, each with a conspicuous clitellum, were placed in plastic boxes (12 × 12 × 8 cm) filled with 300 g of soil, mixed with 3% Mucuna and wetted to field capacity (42%). The boxes were incubated at 26˚C ± 2˚C, and the soil replaced and co- coons collected fortnightly. These cocoons were incubated in Petri dishes at 27˚C. In the greenhouse, plants of M. pruriens were grown for a period of three months (September - Novem- ber 2009). The foliage was then collected, dried (63˚C ± 3˚C, 48 h), ground (to 2 mm), and placed in paper bags; total nitro- gen content of the M. pruriens was determined (2.23%) using the Kjeldahl digestion method following the Mexican Official Norm NOM-021 (SEMARNAT, 2002). Seeds of the studied oak species, Quercus insignis were collected in November 2009 from cloud forest fragments located at 19˚12'N, 96˚59'W, at an elevation of 1460 m . This species bears large acorns (5 cm diameter) and its seeds may be classed as recalcitrant. Seeds were placed in nursery beds (1 × 8 m), where they germinated between 15 - 20 days after sowing, presenting a germination rate > 60%. Experimental Setup The experiment was conducted in the greenhouse of INBI- OTECA from May to November 2010. We used eight treat- ments: 1) Pontoscolex (P); 2) Pontoscolex-Mucuna (PM); 3) Pontoscolex-fertilizer (PF); 4) Pontoscolex-Mucuna-fertilizer (PMF); 5) Mucuna (M); 6) fertilizer (F); 7) Mucuna-fertilizer (MF); 8) Control (C) [only soil]. There were 20 replicates (15 × 25 cm plastic containers) by treatment. The containers were filled with 540 g of dry soil that was then wetted to field capac- ity (43%), and placed on a metallic table. One day later, an 8 - 12 day old Q. insignis seedling of approximate height 15 cm was planted in each container. One week later, two juvenile P. corethrurus earthworms (106 ± 12 mg) were added to the ap- propriate treatments, and Mucuna foliage was added to the soil surface (8 g; 2.23 N%) (nitrogen content of Mucuna is equiva- lent to that of the applied inorganic fertilizer), as well as the slow-release fertilizer (ammonium nitrate [0.7 g; 26 N 13 S%]) according to the requirements of each treatment. Seedlings were grown for a period of 180 days and watered weekly to maintain the soil moisture at field capacity (43%). Mean tem- perature in the greenhouse was 25.6˚C ± 4.3˚C, and weeds were manually removed from the soil. During the experiment, aphids 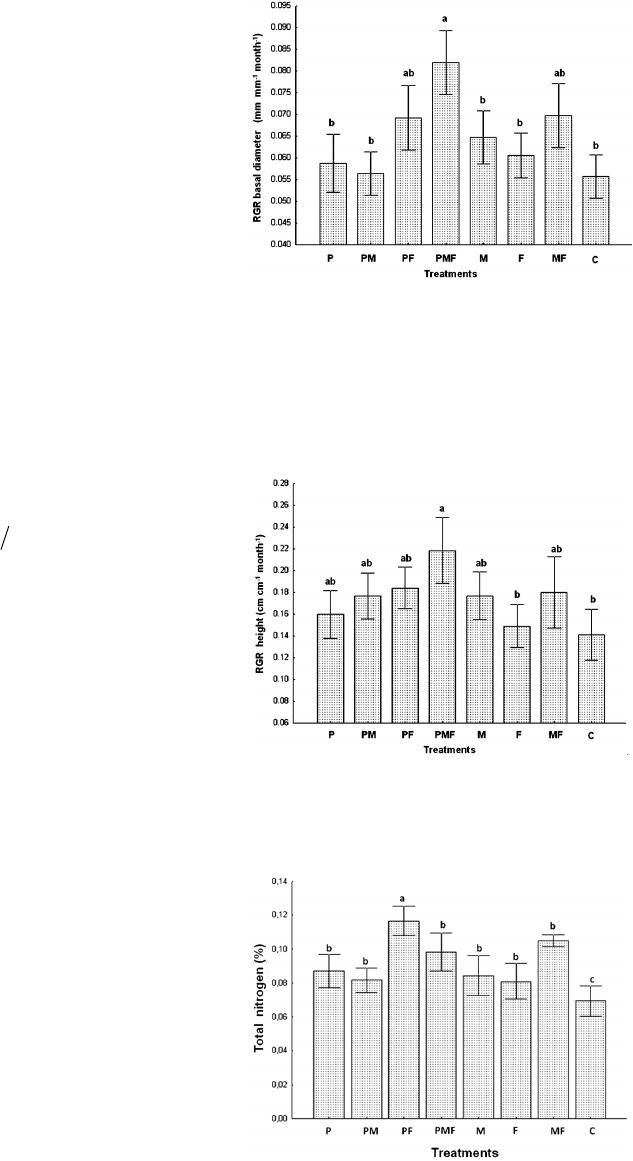 M. L. AVENDAÑO-YÁÑEZ ET AL. OPEN ACCESS were found on the back of some leaves, these were controlled manually and no pesticide was applied. We recorded seedling height and basal diameter monthly. A destructive harvest of all plants from all treatments was carried out 180 days after the start of the experiment. For each plant, the biomass weight was calculated (dried 65˚C ± 3˚C; 72 h) in roots, stem and leaves. To determine leaf nitrogen content a sample of the leaves was taken for each treatment and nitrogen content was determined with the Kjeldahl digestion method, following the Mexican Official Norm NOM-021 (SEMARNAT, 2002). At the end of the study, the number of earthworms and cocoons in each treatment was recorded. Statistical Analysis Analysis of variance (ANOVA) was performed to compare between treatments for each of the following variables: a) growth rate in height, b) growth rate in basal diameter, c) total and component-specific dry biomass, and d) total foliar nitro- gen content. When the ANOVA yielded a significant result, means were compared with a Tukey multiple-comparison test. Prior to the ANOVA, numeric values were transformed to nat- ural logarithm, the normality and homogeneity of variance were fulfilled. Growth rate (TC) was estimated with the following equation: where C2 is final seedling height or diameter, C1 is initial seedling height (cm) or diameter (mm), and t is elapsed time (months). In addition, biomass allocation to leaves (L) was assessed relative to root biomass (R), and expressed as the L/R ratio between treatments. Percentage values of tot al nitroge n con- tent in the leaves were also compared between treatments with a Tukey test (GLM Proc, SAS 9.2; SAS Institute Inc., 2009). Results Growth of Quercus insignis Seedlings Basal diameter growth rate of the Q. insignis was signifi- cantly hi gher in the trea tment PMF (F = 5.33; P < 0.0001) than in all the other treatments (Figure 1). Regarding plant height, significant differences were found (F = 2.84; P < 0.0087) among the treatments PMF, fertilizer (F) and the control treatment (C) (Figure 2). However this trait did not differ significantly between the treatments P. corethru- rus-M. Pruriens-fertilizer (PMF), P. corethrurus-fertilizer (PF) and M. pruriens-fertilizer (MF). Total Leaf Nitrogen Total foliar nitrogen content in Q. insignis seedlings was sig- nificantly different between treatments (F = 2.54; P < 0.01742). Values of this trait were significantly higher in the treatment PF compared to the other treatments. It is noteworthy that the Q. insignis plants that grew in the control treatment (soil only) presented the lowest values of foliar nitrogen, as is shown in Figure 3. Biomass in Q. insignis Seedlings Significant differences between treatments (F = 2.32; P < 0.0290) were recorded for leaf biomass values in Q. insignis. This was particularly noteworthy in the contrast between the P. Figure 1. Effects of eight treatments on growth of Quercus insignis seedlings. Abbreviations: P. corethrurus (P), P. corethru- rus-M. pruriens (PM), P. corethrurus-fertilizer (PF), P. co- rethrurus-M. pruriens-fertilizer (PMF), M. pruriens (M), fertilizer (F), M. pruriens-f ertilizer (MF), Control (C). Mean values of seedling basal diameter are shown. Vertical lines represent standard errors. Different letters indicate signifi- cant differences between treatments. Figure 2. Effects of eight treatments on growth of Quercus insignis seedlings. Mean values of seedling height are shown. Ver- tical lines represent standard errors. Different letters indi- cate significant differences between treatments. Figure 3. Effects of treatments with P. corethrurus (P), M. pruriens (M) and fertilizer (F), and combinations of these, on total leaf nitrogen content in seedlings of Quercus insignis. Values are means f or plants grown in each treat ment. Ver- tical lines represent standard errors. Different letters indi- cate significant differences between treatments.  M. L. AVENDAÑO-YÁÑEZ ET AL. OPEN ACCESS corethrurus-fertilizer (PF) treatment and the control. However the root and shoot biomass Q. insignis seedlings did not differ significantly between treatments (see Table 1). Total biomass was similar between treatments, and signifi- cant differences (F = 3.71; P < 0.0011) were only found among the treatments PF, F and the control (Table 1). The leaf/root biomass ratio was >1 in all treatments, indicating a larger re- source allocation to photosynthetic tissues in Q. insignis seedl- ings, regardless of treatment. No significant differences were observed between treatments in this trait. Discussion It is widely accepted that the presence of earthworms is gen- erally beneficial to plant growth (Edwards & Bohlen, 1996; Bohlen et al., 2002, 2004; Scheu, 2003; Eisenhauer et al., 2009). Similarly, the addition of organic matter to the soil, mediated by leguminous plants such as M. pruriens, is known to provide nutrients such as nitrogen, and to improve soil fertility and thus increase plant productivity (Becker et al., 1995; Konboon et al., 2000). In our study, the treatments with P. corethrurus, M. pruriens, inorganic fertilizer and the interaction between these, presented similar values in terms of growth variables. However, significantly higher values in all plant growth variables were found between P. corethrurus-M. pruriens-fertilizer (PMF) treatment versus the control treatment. Brown et al. (2004) and Scheu (2003) reported increases in the growth of several culti- vated plant species, mostly cereals and grasses, in the presence of earthworms. Likewise, other studies have led to the conclu- sion that the use of Mucuna pruriens favors the growth and productivity of Zea mays (Ile et al., 1996; Buckles & Triomphe, 1999; Eilitta et al., 2003). Our results indicate that the combination of P. corethrurus-M. pruriens-fertilizer (PMF) positively affects the growth in height of Q. insignis plants. However, the combination P. corethru- rus-M. pruriens (PM) and P. corethrurus-fertilizer (PF) can be equally effective in producing increased height, an aspect which is of vital importance to tree species that are propagated in greenhouses. The use of inorganic fertilizers may potentially induce variability in the effects of earthworms on plants, which may largely depend on soil nutrient status (Laossi et al., 2010a). According to our findings, inorganic fertilizer appears to in- teract synergistically with P. corethrurus and M. pruriens, or with M. pruriens only, promoting the growth of Q. insignis during early stages of its life cycle. This condition is likely to be the result of the relatively rapid liberation of nitrogen pro- vided by the synthetic fertilizer. If true, this would partly ex- plain the improved plant growth in the treatments with P. co- rethrurus, M. pruriens and synthetic fertilizer, and in combina- tions of these. The fact that the inorganic fertilizer alone cannot produce a growth increase in these seedlings must not be over- looked. Increases in stem and root biomass in plants that have been exposed to earthworm activity have been repeatedly docu- mented (Wurst & Jones, 2003); however, other studies have reported contrasting results (Scheu, 2003). Ortiz-Ceballos et al., (2007) observed increased root biomass in plants of Zea mays exposed to the combined effects of the activity of the earth- worm Balanteodrilus pearsei with the leguminous M. pruriens. Conversely, the presence of P. corethrurus, and its combination with M. pruriens, did not significantly affect root and stem biomass in Q. insignis seedlings. It must be stressed that most studies reporting increases in root and stem biomass have fo- cused on species with short life cycles (annual or biennial) that are often herbaceous, while the effect of this interaction with long-lived trees is less well understood. Moreover, the effect of earthworms on plant growth may also vary depending on the functional traits of the different plant and earthworm species (Brown et al., 2004; Eisenhauer et al., 2009; Laossi et al., 2010b). As a long-lived tree species, Q. insignis could respond differently in terms of resource allocation to different tissues during a very early phase of its juvenile development. Ulti- mately, this possibility could obscure the synergistic effect of P. corethrurus and M. pruriens on root and shoot biomass alloca- tion in Q. insignis seedlings at the early stages of their devel- opment in a nursery. Leaf biomass is seldom assessed as a response in studies of plant-earthworm interactions. In our analysis, the comparison of leaf biomass of Q. insignis revealed a homogeneous increase between the various treatments, with the exception of the con- trol. The uniform increase of leaf biomass among treatments Table 1. Effects of treatments with P. corethrurus (P), M. pruriens (M) and fertilizer (F), and combination s of these, on the biomass of aboveground and un- derground components and leaf/root ratio of Q. insignis seedlings. Treatments (n) Root biomass (g dry weight) Stem biomass Leaf biomass Total biomass Leaf/root ratio P (17) 2.2 ± 0.9a 2.3 ± 1.2a 3.7 ± 1.7ab 8.3 ± 2.2ab 2.1 ± 1. 2a PM (16) 2.1 ± 0.8a 1.9 ± 0.7a 3.2 ± 1.3ab 7.1 ± 1.5ab 2 ± 1.1a PF (18) 2 ± 0.9a 2.6 ± 1a 4.1 ± 1.3a* 8. 8 ± 2.3a** 2.4 ± 1.6a PMF (16) 1.9 ± 0.9a 1.8 ± 0.8a 3.4 ± 0. 7ab 7 .0 ± 2.1ab 2.2 ± 1.3a M (16) 1.8 ± 0.8a 1.8 ± 1a 2.9 ± 1. 5 ab 6.5 ± 2.2ab 2 ± 1.2a F (17) 1.9 ± 0.6a 1.7 ± 0.9a 2.7 ± 1.4ab 6 .3 ± 2.1b 1.6 ± 0.9a MF (17) 2 ± 0.6a 1.9 ± 0.8a 2.9 ± 1.2ab 6.7 ± 2.5ab 1.7 ± 1. 1a C (17) 1.7 ± 0.4a 1.8 ± 1a 2.6 ± 1.4b 6.1 ± 1.6b 1.6 ± 1a Note: Values are means ± SE. Values in parentheses (n) indicate the number of plants per treatment at the end of the experiment. Different letters within the same column indicate signific ant differences between treatments. *p < 0.0290; **p < 0.0011.  M. L. AVENDAÑO-YÁÑEZ ET AL. OPEN ACCESS involving the addition of P. corethrurus, M. pruriens, the inor- ganic fertilizer, or combinations of these, suggests that plants tend to accumulate photosynthetic tissue when they have unli- mited access to sufficient nutrients provided through biological and/or inorganic fertilization. Moreover, the comparison of total biomass in Q. insignis showed similar patterns among those treatments that combined biological fertilization with inorganic fertilizer, the only significant differences being in the treatment that only included fertilizer, and in the control. Laossi et al. (2010a) reported that the interaction of Lumbricus terre- stris with a fertilizer had significant effects on the total biomass of two herb species, Poa annua and Veronica persica. Similarly, the increase in total biomass of Q. insignis suggests a positive effect of this endogeic earthworm, provided its activity is sup- plemented by an external source of nitrogen that can be assimi- lated by the plant in the short term. Thus, the activity of P. co- rethrurus, combined with the use of M. pruriens and/or chemi- cal fertilizer positively affects both leaf and total biomass in Q. insignis seedling. Total foliar nitrogen content was significantly higher in the treatment P. corethrurus-fertilizer (PF). This result is to be expected given that an increase of leaf nitrogen is related to greater photosynthetic activity, which in theory makes it easier for a plant to access higher quantities of useful resources vital to growth and biomass accumulation in different tissues (Gar- nier, 1991; Gleeson, 1993; Hikosaka, 2004). Although leaf nitrogen concentration often decreases as plants grow (Gastal & Lemaire, 2002), this does not seem to be the case in Q. insignis. This may change, however, later in the life of the plant after its stage transition from sapling to young adult. Plants tend to op- timize resources, and they therefore require a balance in the allocation of nutrients, such as nitrogen, in order to maximize growth (Hilbert, 1990; Gleeson, 1993; Göran & Franklin, 2003). However, the effect of earthworms on this plant resource allo- cation has been poorly documented and understood (Scheu, 2003). In this context, we observed that leaf biomass is gener- ally greater than root biomass in seedlings of Q. insignis, re- gardless of treatment. This situation suggests that the priority for seedlings of this oak species is to produce more photosyn- thetic tissues, which is logical considering that this is a late successional species that establishes under limited light condi- tions in its natural habitat. Conclusion The integration of a biological component such as the earth- worm P. corethrurus into soil management, along with use of M. Pruriens, could favor the growth of Q. insignis. This could be further optimized by the addition of inorganic fertilizer. This combination represents a good option for soil enrichment, di- versification of the practices of plant propagation and reduced use of inorganic fertilizers. Day-to-day practices in nurseries include the application of inorganic fertilizers; however, as this study shows, their use does not necessarily translate into im- proved plant growth. The ultimate goal of biological soil man- agement, including the use of earthworms and green manure, is to promote the integration of edaphic processes such as natural nutrient cycling and degradation to forms that plants can readily assimilate. These processes play decisive roles in the soil dy- namics of an ecosystem and promote robust plant development. Full consideration should therefore be given to the integration of soil biological management into the propagation techniques of native species aimed at conserving and restoring fragmented forest ecosystems. Acknowledgemen ts We thank everyone who helped with this work, Clara Cor- doba-Nieto, Raquel Cervantes-Alday, Álvaro Soberanes and Carolina Cruz. M.L. Avendaño-Yáñez acknowledges the Con- sejo Nacional de Ciencia y Tecnología (CONACYT) for the scholarship granted for this research (Num. Reg. 223896). We also thank the funding received by the Sectorial Fund for Edu- cation Research SEP-CONACYT (project CB-2010-0-156053). REFERENCES Arriaga, V., Cervantes, V., & Vargas-Mena, A. (1994). Manual de reforestación con especies nativas. México: Instituto Nacional de Ecología, Secretaría de Desarrollo Social and Universidad Nacional Autónoma de México. Barois, I., Lav elle, P., Bros sard, M. , Ton doh , J., Mártin ez, M. A., Rossi, J. P., Senapati, B. K., Angeles, A., Fragoso, C., Jiménez, J. J., Decaëns, T., Lattaud, C., Kanyonyo, J., Blanchart, E., Chapuis, L., Brown, G. G., & Moreno, A. (1999). Ecology of earthworm species with large environmental tole rance and/or extended d istributions. In: P. Lavelle, L. Brussard, & P. Hendrix (Eds.), Earthworm manage- ment in tropical agroecosystems (pp. 57-85). Wallingford, UK: CA- BI. Becker, M., Lad ha, J. K., & Ali, M. (1 995). Green manure technolo gy: Potential, usage and li mitations. A case stud y for lowland rice. Plant Soil, 174, 181-194. http://dx.doi.org/10.1007/BF00032246 Benítez, G., Equihua, M ., & Pulido-Salas, M. T. (2002). Diagnóstico d e la situación de los viveros oficiales de Veracruz y su papel para apoyar programas de reforestación y restauración. Revista Chapingo. Serie Ciencias Forestales y del Ambiente, 8, 5-12. Bhadauria, T., & Saxena, K. G. (2010). Role of earthworms in soil fertility maintenance through the production of biogenic structures. Applied and Envi r onmental Soil Science, 2010, Article ID: 816073. http://dx.doi.org/10.1155/2010/816073 Blanchart, E., Villenave, C., Viallatoux, A., Barthes, B., Girardin, C., Azontonde, A., & Feller, C. (2006). Long-term effect of a legume cover crop (Mucuna pruriens var. utilis) on the communities of soil macrofauna and nematofauna, under maize cultivation, in southern Benin. European Journal of Soil Biology, 42, 136-144. http://dx.doi.org/10.1016/j.ejsobi.2006.07.018 Bohlen, P. J., Edwards, C.A., Zhang, Q., Parmelee, R. W., & Allen, M. (2002). Indirect effects of earthworms on microbial assimilation of labile carbon. Applied Soil Ecology, 20, 255-261. http://dx.doi.org/10.1016/S0929-1393(02)00027-6 Bohlen, P. J., Parmelee, R. W., & Blair, J. M. (2004). Integrating the effects of earthworms on nutrient cyclin g across spatial and temporal scales. In: C. A. Edwards (Ed.), Earthworm ecology (pp. 161-180). Boca Raton, FL: CRC Press. http://dx.doi.org/10.1201/9781420039719.ch9 Brown, G. G., Pashanasi, B., Villenave, C., Patron, J. C., Senapati, B. K., Giri, S,, Barois, I., Lavelle, P., Blanchart, E., Blakemore, R. J., Spain, A. V., & Boyer, J. (1999). Effects of earthworms on plant production in the tropics. In: P. Lavelle, L. Brussard, & P. Hendrix (Eds.), Earthworm management in tropical agroecosystems (pp. 87- 137). Wallingford, UK: CABI. Brown, G. G., Edwards, C. A., & Brussaard, L. (2004). How earth- worms affect plant growth: b urrowing in to the mechanisms. In: C. A. Edwards (Ed.), Earthworm ecology (pp. 13-49). Boca Raton, FL: CRC Press. Buch, A. C., Brown, G. G., Niva, C. C., Sautterc , K. D., & Lourenc, L. F. (2011). Life cycle of Pontoscolex corethrurus (Müller, 1857) in tropical artificial soil. Pedobiologia, 54, S19-S25. http://dx.doi.org/10.1016/j.pedobi.2011.07.007 Buckles, D. (1995). Velvetb ean: A new plan t with a history. Economic  M. L. AVENDAÑO-YÁÑEZ ET AL. OPEN ACCESS Botany, 49, 151-162. http://dx.doi.org/10.1007/BF02862271 Buckles, D., & Triomphe, B. (1999). Adoption of mucuna in the farm- ing system s of northern Honduras. Agroforestry Systems, 47, 67-91. http://dx.doi.org/10.1023/A:1006205702691 Cayuela, L., Golich er, D. J., Benayas, J. M. R., González-Espino sa, M., & Ramírez-Marcial, N. (2006). Fragmentation, disturbance and tree diversity conservation in tro pical mon tan e f orests . Journal of Applied Ecology, 43, 1172-1181. http://dx.doi.org/10.1111/j.1365-2664.2006.01217.x Doube, B. M., Williams, P. M. L., & Willmott, P. J. (1997). The influ- ence of two species of earthworm (Aporrectodea trapezoids and Aporrectoedea rosea) on the growth of wheat, barley and faba beans in three soil types in the greenhouse. Soil Biology and Biochemistry, 29, 503-509. http://dx.doi.org/10.1016/S0038-0717(96)00037-5 Edwards, C. A., & Bater, J. E. (1992). The use of earthworms in envi- ronmental management. Soil Biology and Biochemistry, 24, 1683- 1689. http://dx.doi.org/10.1016/0038-0717(92)90170-3 Edwards, C. A., & Bohlen, P. (1996). Biology and ecology of earth- worms. New York: Chapman and Hall. Eisenhauer, N., Milcu, A., Sabais, A. C. W., & Scheu, S. (200 9). Earth - worms enhance plant regrowth in a grassland plant diversity gradient. European Journal of Soil Biology, 45, 455-458. http://dx.doi.org/10.1016/j.ejsobi.2009.06.001 Eilitta, M., Sollenberger, L. E., Littell, R. C., & Harrington, L. W. (2003). On-farm experiments with maize-mucuna systems in the Los Tuxtlas region of Veracruz, Mexico. I. Mucuna biomass and maize grain yield. Experimental Agriculture, 39, 5-17. http://dx.doi.org/10.1017/S0014479702001126 Fragoso, C., Kanyonyo, J., Moreno, A., Senapati, B., Blanchart, E., & Rodríguez, C. (1999). A survey of tropical earthworms: Taxonomy, biogeography and environmental plasticity. In: P. Lavelle, L. Brus- sard, & P. Hendrix (Eds.), Earthworm management in tropical agroecosystems (pp. 1-26). Wallingford, UK: CABI. Fragoso, C. (2001). Las lombrices de tierra de México (Annelida, Oligochaeta): Diversidad, ecología y manejo. Acta Zoológica Mex- icana, 1, 131-171. Garnier, E. (1991). Reso urce capture, bio mass allocation and growth in herbaceous plants. Trends in Ecology & Evolution, 6, 126-131. http://dx.doi.org/10.1016/0169-5347(91)90091-B Gastal, F., & Lemaire, G. (2002). N uptake and distribution in crops: An agronomical and ecophysiological perspective. Journal of Expe- rimental Botany, 53, 789-799. http://dx.doi.org/10.1093/jexbot/53.370.789 Gleeson, S. K. (1993). Optimization of tissue nitrogen and root-shoot allocation. Annals of Botany, 71, 23-31. http://dx.doi.org/10.1006/anbo.1993.1003 González-Espinosa, M., Meave, J. A., Lorea-Hernández, F. G., Ibarra- Manríquez, G., & Newton, A. C. (2011). The red list of Mexican cloud forest trees. Cambridge: Fauna and Flora International (FFI). Göran, I. A., & Franklin, O. (2003). Root: Shoot ratios, optimization and nitrogen productivity. Annals of Botany, 92, 795-800. http://dx.doi.org/10.1093/aob/mcg203 Haimi, J., & Einbork, M. (1992). Effects of endogeic earthworms on soil processes and plan t growth in con iferous forest so il. Biology and Fertility of Soils, 13, 6-10. http://dx.doi.org/10.1007/BF00337230 Hilbert, D. W. (1990). Optimization of plant root: Shoot ratios and internal nitrogen concentration. Annals of Botany, 66, 91-99. Hikosaka, K. (2004). Interspecific difference in the photosynthesis- nitrogen relationship: Patterns, physiological causes, and ecological importance. Journal of Plant Research, 6, 481-494. http://dx.doi.org/10.1007/s10265-004-0174-2 Hulugalle, N. R., Lail, R., & Kuile, C. H. H. (1986). Amelioration of soil physical properties by Mucu na after mechan ized land clearing o f tropical rain forest. Soil Science, 14, 219-224. http://dx.doi.org/10.1097/00010694-198603000-00007 Ile, E., Hamadin a, M. K., Zufa , K., & Henrot, J. (1996 ). Note on effects of a Mucuna pruriens var. utilis crop on the growth of maize (Zea mays) on an acid ultisol in southeastern Nigeria. Field Crops Re- search, 48, 135-140. http://dx.doi.org/10.1016/S0378-4290(96)01028-3 Konboon, Y., Blair, G. J., Lefroy, R. D. B., & Whitbread, A. M. (2000). Tracing the nitrogen, sulfur and carbon released from plant residues in a soil/plant system. Australian Journal of Soil Research, 38, 699- 710. http://dx.doi.org/10.1071/SR99075 Lamb, D., Erskine, P. D., & Parrota, J. A. (2005). Restoration of de- graded tropical forests landscapes. Science, 310, 1628-1632. http://dx.doi.org/10.1126/science.1111773 Laossi, K. P., Ginot, A., Noguera, D. C., Blouin, M., & Barot, S. (2010a). Earthworm effects on plant growth do not necessarily de- crease with soil fertility. Plant and Soil, 328, 109-118. http://dx.doi.org/10.1007/s11104-009-0086-y Laossi, K. P., Noguera, D. C., & Barot, S. (2010b). Earthworm me- diated maternal effects on seed germination and seedling growth in three annual plants. Soil Biology and Biochemistry, 42, 319-323. http://dx.doi.org/10.1016/j.soilbio.2009.11.010 Lavelle, P., Ba rois, I., Cruz, I. , Fragoso, C., He rnández, A., Pin eda, A., & Rangel, P. (1987). Adaptative strategies of Pontoscolex corethru- rus (Glossoscolecidae, Oligochaeta), a peregrine geophagous earth- worm of the hum id tropics. Biology and Fertility of Soils, 5, 188-194. http://dx.doi.org/10.1007/BF00256899 Lavelle, P., & Pashanasi, B. (1989). Soil macrofauna and land man- agement in Peruvian A mazonia (Yurimaguas, Loreto). Pedobiologia, 33, 283-291. Lee, K. E. (1985). Earthworms: Their ecology and relationships with soils and land use. Sydney: Academic Press. López-Hernández, D., Lavelle, P., Fardeau, J. C., & Niño, M. (1993). Phosphorus transformations in two P-sorption contrasting tropical soils during transit through Pontoscolex corethrurus (Glossoscoleci- dae: Oligochaeta). Soil Biology and Biochemistry, 25, 789-792. http://dx.doi.org/10.1016/0038-0717(93)90124-T Meli, P. (2003). Restauración ecológica de bosques tropicales: Veinte años de investigación académica. Interciencia, 28, 581-589. Meza-Sánchez, R., Ruiz-Espin oza, F. H., & Navejas-Jiménez, J. (2009). Guía para la producc ión de planta y plantaci ón con especies nativas. Baja California Sur: Instituto Nacio nal de Investig aciones Forestales, Agrícolas y Pecuarias (INIFAP). Müller, F. (185 7). II. Description of a new species of earthwo rm (Lum- bricus corethrurus). Journal of Natural History Series 2, 20, 13-15. http://dx.doi.org/10.1080/00222935709487865 Oldfield, S., & Eastwood, A. (2007). The red list of oaks. Cambridge: Fauna and Flora International (FFI). Ortiz-Ceballos, A. I., & Fragoso, C. (2004). Earthworm populations under tropical maize cultivation: The effect of mulching with vel- vetbean. Biology and Fertility of Soils, 39, 438-445. http://dx.doi.org/10.1007/s00374-004-0732-8 Ortiz-Ceballos, A. I., Frago so, C ., Eq u ihu a, M. , & Bro wn , G. G. (2005). Influence of food quality, soil moisture and the earthworm Pontos- colex corethrurus on the growth, reproduction and activity of a trop- ical earthworm Balanteodrilus pearsei. Pedobiologia, 49, 89-98. http://dx.doi.org/10.1016/j.pedobi.2004.08.006 Ortiz-Ceballos, A. I., Frag oso, C., & Brown, G. G. (2007). Synergistic effect of a tropical earthworm Balanteodrilus pearsei and velvetbean Mucuna pruriens var. utilis on maize growth and crop production. Applied Soil E cology, 35, 356-362. http://dx.doi.org/10.1016/j.apsoil.2006.07.009 Ortiz-Ceballos, A. I., Aguirre-Rivera, J. R., Osorio-Arce, M. M., & Peña-Valdivia, C. (201 2). Velvet Bean (Mucuna pruriens var. utilis) a cover crop as Bioherbicide to preserve the environmental services of soil. In: R. Alvarez-Fernandez (Ed.), Herbicides-environmental impact studies and management approaches (pp. 167-184). Cam- bridge: University of Cambridge. Pashanasi, B., Meléndez, G., Szott, L., & Lavelle, P. (1992). Effect of inoculation with the endogeic earthworm Pontoscolex corethrurus (Glossoscolecidae) on availability, soil microbial biomass and the growth of three tropical f ruit tree seedlings in a pot experiment. Soil Biology and Biochemistry, 24, 1655-1659. http://dx.doi.org/10.1016/0038-0717(92)90165-T Pedraza, R. A., & Williams-Linera, G. (2003). Evaluat ion o f nativ e tree species for the rehabilitation of deforested areas in a Mexican cloud forest. New Forests, 26, 83-99.  M. L. AVENDAÑO-YÁÑEZ ET AL. OPEN ACCESS http://dx.doi.org/10.1023/A:1024423511760 Ramírez-Marcial, N., González-Espinosa, M., & Williams-Linera, G. (2001). Anthropogenic d isturbance a nd tree diversit y in montane rain forests in Chiapas, Mexico. Forest Ecology and Management, 154, 311-326. http://dx.doi.org/10.1016/S0378-1127(00)00639-3 Saunders, D. A., Hobbs, R. J., & Margules, C. R. (1991). Biological consequences of ecosystem fragmentation: A review. Conservation Biology, 5, 18-32. http://dx.doi.org/10.1111/j.1523-1739.1991.tb00384.x Senapati, B. K., Lavelle, P., Giri, S., Pashanasi, B., Alegre, J., Decaëns , T., Jimenez, J. J., Albrecht, A., Blanchart, E., Mahieux, M., Rous- seaux, L., Thomas, R., Panigrahi, P. K., & Venkatachalam, M. (1999). In-soil earthworm technologies for tropical agroecosystems. In: P. Lavelle, L. Brussaard, & P. Hendrix (Eds.), Earthworm man- agement in tropical agr oecosystems (pp. 199-237). Wallingford, UK: CABI. SAS Institute Inc. (2009). SAS system for windows version 9.2. http://www.sas.com/ SEMARNAT (2002). Nom -021-SEMARNAT-2000 Que establece las especificaciones de fertilidad, salinidad y clasificación de suelos, estudio, muestreo y análisis, 2nd Sect. México : Secretaría del Medio Ambiente y Recursos Naturales (SEMARNAT). Scheu, S. (2003). Effects of earthworms on plant growth: Patterns and perspectives. Pedobiologia, 47, 846-856. http://dx.doi.org/10.1078/0031-4056-00270 Smyth, T. J., Cravo, M. S., & Melgar, R. J. (1991). Nitrogen supplied to corn by legumes in a Central Amazon Oxisol. Tropical Agriculture (Trinidad), 68, 366-372. Valencia, A. S. (2004). Diversidad del género Quercus (Fagaceae) en México. Boletin de la Sociedad Botanica de México, 75, 33-53. Vázquez-Yanes, C., & Cervantes, V. (199 3). Reforestación con árboles nativos de México. Ciencia y Desarrollo, 19, 52-58. Villaseñor, J. L. (2 0 1 0 ). El bosque húmedo de montaña en México y sus plantas vasculares: Catálogo florístico-taxonómico. México: Comi- sión Nacional para el Conocimiento y Uso de la Biodiversidad (CONAB IO) —Universidad Nacional Autónoma de México (UNAM). Wardle, D. A. (2002). Communities and ecosystems: Linking above- ground and belowground components. Princeton, NJ: P rinceton Uni- versity Press. Wardle, D. A., Bardgett, R. D., Kironomos, J. N., Setälä, H., Van der Putten, W. H., & Wall, D. H. (2004). Ecological linkages between aboveground and belowground biota. Science, 304, 1629-1633. http://dx.doi.org/10.1126/science.1094875 Wurst, S., & Jon es, T. H. (2003 ). Indirect eff ects of earthworms (Apor- rectodea caliginosa) on an above-ground tritrophic interaction. Pe- dobiologia, 47, 91-97. http://dx.doi.org/10.1078/0031-4056-00173
|