1. Introduction
Myracrodruon urundeuva (Allemão) Engl., Anacardiaceae, popularly known as aroeira, backwoods aroeira or urundeuva, is a deciduous species, heliophytic and selective xerofita (Lorenzi, 1992) . M. urundeuva occurs naturally in the extreme northwest of Argentina, in the south and east of Bolivia and Paraguay. In Brazil, it occurs in the following states: Paraíba, Pernambuco, Piauí, Rio de Janeiro, Rio Grande do Norte, Sergipe, São Paulo, Tocantins, Federation District and Mato Grosso do Sul (Carvalho, 2003) . It occurs also in different phytoecological regions, as semi deciduous forest; deciduous forest; scrubland and savanna; caatinga; kills drought; Mato Grosso do Sul and Mato Grosso Pantanal Chaco (Carvalho, 1994) and forest mono-dominated in Minas Gerais (Oliveira et al., 2015) . The wide geographic distribution of a species is an indicator of high levels of genetic diversity, which can give it the ability to occupy different habitats (Kageyama et al., 2003) .
The species has high-density wood, excellent mechanical performance and good chemical and physical defense, which explains the strength, hardness and durability. It is considered the toughest wood from Brazil. While one square centimeter concrete supports a load of 250 kgf wood, M. urundeuva can support 696 kgf, and it is registered at the Institute for Technological Research (IPT) in São Paulo, such as wood “durable” and put in select group of woods with secular durability, calls “imputrescible” (Ribeiro, 1989) . Moreover, M. urundeuva was identified as the most potential species for research on phytochemicals (Pereira et al. 2014) . Whereas M. urundeuva is endangered (BRASIL, 2008) , Souza et al. (2012) reported a significant use of this species for timber purposes, due to the popularity and the timber and non-timber value of M. urundeuva, can lead to local extinction. Due to these facts, M. urundeuva is experiencing a strong exploration process, in a predatory way, causing the devastation of their natural populations. In addition, it should be noted that selective logging for use in the timber industry virtually wiped out the large individuals, therefore considered endangered and categorized as vulnerable (Brandão, 2000; Mendonça & Lins, 2000) .
Studies to preserve and guarantee the genetic variability of these threatened populations have been intensified, and the tools used are the progeny tests, which is an ex situ strategy. Currently one of the major purposes of the progeny test is its transformation into an orchard, to provide seeds with genetic quality to restore the Legal Forest with superior material and adapted soil and climatic conditions associated with economic alternatives, to ensure the population’s quality of life place. Thus, the aim of this study was to evaluate the genetic variability in nine M. urundeuva progeny tests.
2. Materials and Methods
Seeds of six natural populations of M. urundeuva were collected in two situations, without human disturbance (natural forest) and with human disturbance (modified areas with agriculture and grazing) (Table 1). Which were seedlings produced and installed in nine progeny tests in Education, Research and Extension Farm, located in Selvíria, Mato Grosso do Sul State, Brazil, thus, constituting the M. urundeuva ex situ bank.
With the seeds from natural populations of M. urundeuva without human disturbance of Paulo de Faria, four progeny tests were installed. The other populations been installed in only one provenance and progeny test. All experiments were conducted, using the randomized block design. The number of replications; the number of progenies and the number of plants per plot are presented in Table 2.
![]()
Table 1. Geographical Location of natural populations of Myracrodruon urundeuva with human disturbance (CPA) and without human disturbance (SPA).
![]()
Table 2. Characteristics of Progeny Tests (TP) of Myracrodruon urundeuva installed in Selvíria, Mato Grosso do Sul.
R: number of replications; P: number of progeny; N: number of plants by progenies.
In the test 1, 3, 4, 5, 8 and 9, M. urundeuva was planted in consortium with other species: 1-Anadenthera falcata and Guazuma ulmifolia; 3 and 4-Ricinus communis, Zea mays and Cajanus cajan; 5-Eucalyptus spp; 8-Trema micranta, Peltophorum dubium and Eucalyptus spp, 9-Cordia trichotoma, because M. urundeuva needs shade in early development. The progeny tests 2, 6 and 7 were planted only using M. urundeuva. All trials were measured in 2007 for the quantitative traits: total tree height (H); average crown diameter (DMC) and diameter at breast height (DBH).
Quantitative variables were analyzed by the method of linear mixed model (univariate additive)-REML/BLUP, applied to progeny tests in randomized block design, many plants per plot, one site and a single population, following the procedure proposed by Resende (2002a) : y = Xb + Za + Wc + e; where: y = data vectors; b = vector of the effects of blocks (fixed); a = vector of additive genetic effects (random); c = vectors of the effects of plot (random); e = vector of random errors effects. X, Z e W = incidence matrices for b, a and c, respectively. Estimates of variance components and genetic parameters were obtained by REML/BLUP method (restricted maximum/best linear prediction likelihood untainted), using genetic-statistical software SELEGEN- REML/BLUP (Resende, 2002b) . The estimated parameters were the coefficients: heritability in individual, strict sensus (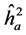 ), average among progenies (
), average among progenies (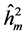 ) and additive within plot (
) and additive within plot (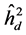 ); genetic variation at the individual level (CVgi) and plot level (CVgp); relative variation (CVr) and accuracy (Ac) to the traits H, DMC and DBH.
); genetic variation at the individual level (CVgi) and plot level (CVgp); relative variation (CVr) and accuracy (Ac) to the traits H, DMC and DBH.
3. Results and Discussion
The highest average annual increment (IMA) for the height (Table 3) for populations without human disturbance was found in the TP-3 (0.89 m) and the lowest in TP-4 (0.58 m), and for the population with human disturbance, the highest was in TP-9 (1.20 m) and the lowest was in TP-8 (0.43 m). Sebbenn and Etorri (2001) , studying a progeny
![]()
Table 3. Average estimates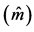 , mean annual increment (IMA), experimental variation coefficient (CVexp), change index (IV), F-test (F) and correlation due to the common environment of the plot
, mean annual increment (IMA), experimental variation coefficient (CVexp), change index (IV), F-test (F) and correlation due to the common environment of the plot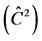 , for traits height (H), DMC and DBH.
, for traits height (H), DMC and DBH.
test of M. urundeuva consortium with Peltophorum dubium and Esenbechia leiocarpa found IMA H of 0.89 m. Bertoni and Dickfeldt (2007) evaluating M. urundeuva planted in alternating areas in the State Park of Porto Ferreira, Sao Paulo State, Brazil, found for H an IMA 0.72 m.
In general, environmental control was good for all trials (CVexp < 20%, Pimentel Gomes (1985) . But the variation index (IV) showed a lower percentage because the value depends only on CVexp, as a proportion of the residual variation of experimental medium (Pimentel Gomes, 1987) and the variation index proposed by Pimentel Gomes (1991) or experimental precision coefficient Storck et al. (2000) consider the number of replications used in the trial, apart from a residual nature variation.
The TP-4 showed significant difference at 1% of probability among progenies, indicating genetic variation among the progeny for the traits height; in other progeny tests occurred homogeneity of values, which was acquired with the development of the progeny, as in earlier ages had differences. When the TP-8 and the TP-3 were evaluated at younger ages, Fonseca (2003) ; Freitas, Moraes and Buzetti (2002) and Freitas et al. (2006) detected significant differences at 5% of probability for height.
Correlation due to the common environment of the plot 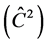 in TP-4 and TP-8 showed to be the most suitable trials for the selection for height, because there is little environmental influence on the variation observed in the plots. According to Sturion and Resende (2005) , the values of around 0.10 are observed in experiments with perennials plants.
in TP-4 and TP-8 showed to be the most suitable trials for the selection for height, because there is little environmental influence on the variation observed in the plots. According to Sturion and Resende (2005) , the values of around 0.10 are observed in experiments with perennials plants.
The highest means of DMC were found in the TP-5 and TP-7, the CVexp ranged from 8.4 to 22.8%, which shows appropriate experimental environmental control in the trials. The variation rates (IV), which takes into account the number of replications, ranged from 0.70 to 7.60, these values can be considered low for experiments where there is competition (Pimentel Gomes and Garcia, 2002) .
The F test indicated that there is genetic variability among progenies in TP-4, 6 and 8 for the DMC, with significant differences at the level of 1% and 5%. The TP-2 and 6 showed lowest value for the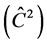 , 0.07 and 0.03, respectively. The other progeny tests presented values higher than 10%.
, 0.07 and 0.03, respectively. The other progeny tests presented values higher than 10%.
For DBH the highest averages were found in TP-6 (7 cm) and 7 (12.94 cm). The highest IMA value was 0.85 cm for TP-2/ These values are superior to that found by Guerra et al. (2009) , obtained average DBH of 6.87 cm and 5.98 cm for M. urundeuva originating from Aramina-SP and Selvíria-MS, at 15 years of age.
The CVexp ranged from medium to high, but the IV indicated good environmental control. TP-4, 5 and 6 showed significant differences between progenies for DBH. The correlation due to the common environment of the plot revealed that the DBH is the most favorable trait for selecting.
The average heritability among progenies (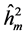 ) was greater than the individual heritability (
) was greater than the individual heritability (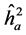 ) and additive within plots (
) and additive within plots (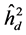 ) for all trait in all of progeny tests (Table 4). This results are in agreement with the results found by Siqueira et al. (2000) , in Balfourodendron riedelianum, Sebbenn and Ettori (2001) in M. urundeuva, Sebbenn et al. (2003) in Araucaria angustifolia, Souza et al. (2003) in Astronium fraxinifolium, Costa et al. (2005) in Ilex paraguariensis, Moraes et al. (2007) in Hevea brasiliensis,
) for all trait in all of progeny tests (Table 4). This results are in agreement with the results found by Siqueira et al. (2000) , in Balfourodendron riedelianum, Sebbenn and Ettori (2001) in M. urundeuva, Sebbenn et al. (2003) in Araucaria angustifolia, Souza et al. (2003) in Astronium fraxinifolium, Costa et al. (2005) in Ilex paraguariensis, Moraes et al. (2007) in Hevea brasiliensis,
![]()
Table 4. Genetic parameter estimates for the traits height, DMC and DBH for testing progenies Myracrodruon urundeuva with and without human disturbance.
Missio et al. (2005) in Pinus caribaea var. hondurensis, and Oliveira et al. (2004) in Spondias tuberosa.
The 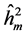 was higher in progeny tests originated from areas with human disturbance, except for the DBH. This results for all the traits studied indicate that the selection can be more effective by selection among progenies (Costa et al., 2010) . According to Falconer (1987) , small populations are expected to present lower heritability than large populations. The average heritability among progeny show great generic control of traits, indicating the possibility to obtain genetic gains by selecting the best families for seed production.
was higher in progeny tests originated from areas with human disturbance, except for the DBH. This results for all the traits studied indicate that the selection can be more effective by selection among progenies (Costa et al., 2010) . According to Falconer (1987) , small populations are expected to present lower heritability than large populations. The average heritability among progeny show great generic control of traits, indicating the possibility to obtain genetic gains by selecting the best families for seed production.
The highest heritability coefficients for height were obtained in TP-4 (Table 3). The TP-4 presented highest heritabilities in the first yea (0.8, 0.36 and 0.07, for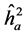 ,
, 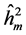 ,
, 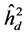 , respectively) and in the second year (0.35, 0.75 and 0.28 to
, respectively) and in the second year (0.35, 0.75 and 0.28 to![]() ,
, ![]() ,
, ![]() , respectively). According Ettori et al. (2006) the coefficient of heritability shows variation with age of the plant, because until there is full to the countryside and balance, there may be greater or lesser influence of the environment on the expression of growth traits.
, respectively). According Ettori et al. (2006) the coefficient of heritability shows variation with age of the plant, because until there is full to the countryside and balance, there may be greater or lesser influence of the environment on the expression of growth traits.
In progeny tests derived from the Ecological Station of Paulo de Faria (TP-1, 2, 3 and 5) the![]() ) ranged from 0.03 to 0.55 for height, 0.01 to 0.48 for DMC and, 0.03 to 0.53 for DBH. Such variations show that heritability is a property not only of the traits, but also the population and environmental circumstances to which the progenies are subject of investigations (Falconer, 1987) .
) ranged from 0.03 to 0.55 for height, 0.01 to 0.48 for DMC and, 0.03 to 0.53 for DBH. Such variations show that heritability is a property not only of the traits, but also the population and environmental circumstances to which the progenies are subject of investigations (Falconer, 1987) .
The presence of genetic variability can be confirmed and quantified by the coefficient of genetic variation, which expresses the magnitude of genetic variation relative to the nature of the medium (Resende et al., 1991) . The greatest genetic variation values were found in the DMC and DBH in TP-4 (17.3 and 2.8, respectively), indicating that there is a greater amount of genetic variation among progenies from areas without human disturbance. Incontrast, occurred less genetic variation when compared to populations from forest fragments.
The coefficient of genetic variation in individual level (CVgi) was in general twice time large than the CVgp (Table 3) for height, DMC and DBH. According to Aguiar et al. (2010) , the larger value for the greater ease to find individuals that may provide higher gains in the selection.
The CVr indicates the most appropriate trait for selection, and the TP-5, 7 and 9 the height, the TP-2, 3, 6, 8 and 9 the DMC, the TP-1 and 4 DBH. According Vencovsky and Barriga (1992) , traits that have values close to one are suitable for carrying out a selection.
The accuracy, which measures the confidence level of the estimates, was high in TP-4 for DBH, in TP-4, 5 and 7 for height and TP-4, 6 and 8 for the DMC, according with the magnitude criterion of Resende and Duarte (2007) . Thus, the M. urundeuva progeny tests provide genetic variability among progenies enough to conduct genetic conservation and breeding programs.
4. Conclusion
The evaluations of silvicultural traits in the nine M. urundeuva progeny tests allowed the observation that the trees had a satisfactory development.
Genetic variation of the populations with human disturbance is lower than populations originated from areas without human disturbance.
The progeny tests showed a high accuracy, which ensures the viability of the genetic conservation bank ex situ and success in the breeding programs, by the transformation of trials seed orchards. The most appropriate trait for selection is the diameter at breast height.
Acknowledgements
To the Pro-Rectory of Research for the Young Investigator grant. To aid the search FAPESP No. 05/60113-7 and CNPq No. 470713/2006-2. CNPq for the PhD scholarship. The TT2 FAPESP Grant No. 06/55191-1. The technical agricultural João Antonio de Souza Queiroz and employees of UNESP Jose Cambuim and Alonso Angelo da Silva for the help in the development of this research.