Permanence, Periodicity and Extinction of a Delayed Biological System with Stage-Structured Preference for Predator ()
Received 27 January 2016; accepted 26 March 2016; published 29 March 2016

1. Introduction
In recent years, much attention has been paid to biological systems with stage structure [1] - [23] . One important reason is that there are many species whose individual members have a life history taking them through two stages, immature and mature. Thus considering stage structure in population corresponds with the natural phenomenon. Another reason is that stage-structured ecological models are much simpler than the models governed by partial differential equations but they can exhibit phenomena similar to those of partial differential equations and many important physiological parameters can be incorporated [24] . The other reason is that the biological dynamics has long been and will continue to be one of the dominant themes in both ecology and mathematical ecology due to its universal existence and importance [25] .
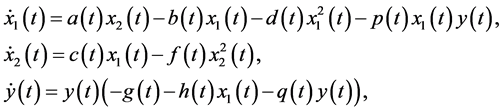 (1.1)
(1.1)
where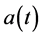 ,
, 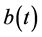 ,
, 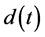 ,
, 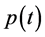 ,
, 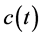 ,
,  ,
, 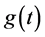 ,
,  and
and 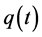 are all continuous positive T-periodic functions, and
are all continuous positive T-periodic functions, and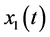 ,
, 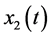 , and
, and 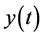 denote the densities of immature prey, mature prey, and predator species, respectively. They obtained a set of sufficient and necessary conditions that guarantee the permanence of the system.
denote the densities of immature prey, mature prey, and predator species, respectively. They obtained a set of sufficient and necessary conditions that guarantee the permanence of the system.
In the natural world, many predators switch to alternative prey when their favored food is in short supply [22] - [24] . For example, the lynx switches to red squirrel when the snowshoe hare is scarce [25] . Even if there is only one prey type, the degree of predation or the quality (including palatability) of prey is likely to vary with its stage structure, which is likely to affect the predator’s preference for different stage-structured prey. This type of behavior has been reported in Asecodes hispinarum [26] , who parasitizes all 5 instars of Brontispa Logissina, but prefers to parasitize the 2nd and 3rd instars when it is exposed to all the instars of larvae, and in Microplitis mediator [27] , who prefers to parasitize the 2nd and 3rd instars of Mythimna separate.
However, previous studies on prey age preference only have been done in laboratory tests. Few researchers have investigated the phenomenon with mathematical models and carried out theoretical analysis together with numerical simulation. To extend research in this area, and based on the recent study by Cui and Song [21] , we consider a periodic predator-prey system with time delay and a predator with stage-structured preference.
2. Formulation of the Model
Let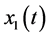 ,
, 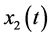 and
and ![]() represent the density of immature prey, mature prey and predator species, respectively. Our periodic predator-prey system with time delay and stage-structured preference of the predator can be described as following:
represent the density of immature prey, mature prey and predator species, respectively. Our periodic predator-prey system with time delay and stage-structured preference of the predator can be described as following:
![]() (2.1)
(2.1)
where
![]() (2.2)
(2.2)
The coefficients in system (2.1) are all continuous positive T-periodic functions. Parameter ![]() is the immature prey preference of the predator, which takes a value between 0 and 1;
is the immature prey preference of the predator, which takes a value between 0 and 1; ![]() is the mature prey preference of the predator [28] [29] .
is the mature prey preference of the predator [28] [29] . ![]() (
(![]() ) denotes the predation weighting factor for
) denotes the predation weighting factor for![]() . The parameters
. The parameters ![]() and
and ![]() represent the birth rate and the death rate of the immature prey populations, respectively.
represent the birth rate and the death rate of the immature prey populations, respectively.
The term ![]() represents the numbers of immature prey born at time
represents the numbers of immature prey born at time ![]() that still
that still
survive, and those progressing from the immature stage to the mature stage at time t. The death rate of the mature prey population is logistic in nature and it is proportional to the square of the population with proportionality![]() . The parameter
. The parameter ![]() and
and ![]() denote the death rate and the overcrowding rate or overcrowding effecting of the predator population, respectively. The overcrowding effecting
denote the death rate and the overcrowding rate or overcrowding effecting of the predator population, respectively. The overcrowding effecting ![]() denote the phenomenon that population growth rate is decreased with the increase of density. The function
denote the phenomenon that population growth rate is decreased with the increase of density. The function ![]() represents the Beddington-DeAngelis functional response of the predator to the immature prey and
represents the Beddington-DeAngelis functional response of the predator to the immature prey and ![]() is the conversion rate of nutrients into the reproduction of the predator. The parameter
is the conversion rate of nutrients into the reproduction of the predator. The parameter ![]() is the delay due to gestation, that is to say, only the mature adult predator can contribute to the production of predator biomass. The functional response of the predator to the mature prey takes the Holling type-III form of
is the delay due to gestation, that is to say, only the mature adult predator can contribute to the production of predator biomass. The functional response of the predator to the mature prey takes the Holling type-III form of ![]() and
and ![]() denotes the conversion rate of nutrients into the reproduction of the predator.
denotes the conversion rate of nutrients into the reproduction of the predator.
The initial conditions for system (2.1) take the form of
![]() (2.3)
(2.3)
where![]() ,
, ![]() , the Banach space of continuous functions
, the Banach space of continuous functions
mapping the interval ![]() into
into![]() , where we define
, where we define ![]() and the interior of
and the interior of
![]() as
as![]() .
.
For continuity of initial conditions, we require
![]() . (2.4)
. (2.4)
For the purpose of convenience, we write
![]() .
.
Obviously, ![]() is a T-periodic and strictly positive function. Then system (2.1) becomes
is a T-periodic and strictly positive function. Then system (2.1) becomes
![]() (2.5)
(2.5)
In this paper, we consider system (2.5) with initial conditions (2.3) and (2.4). At the same time, we adopt the following notation through this paper:
![]() ,
, ![]() ,
, ![]() ,
,
where ![]() is a continuous T-periodic function.
is a continuous T-periodic function.
The rest of the paper is arranged as follows. In the following section, we introduce some lemmas and then explore the permanence and periodicity of system (2.5). In Section 4, we investigate the extinction of the predator population in system (2.5). In Section 5, numerical simulations are presented to illustrate the feasibility of our main results. Furthermore, the simulated results are explained according to the biological perspective. In section 6, a brief discussion is given to conclude this work.
3. Permanence and Periodicity
In this section, we analyze the permanence and periodicity of system (2.5) with initial conditions (2.3) and (2.4). Firstly, we introduce the following definition and Lemmas which are useful to obtain our result.
Definition 3.1. The system![]() ,
, ![]() is said to be permanent if there are constants
is said to be permanent if there are constants ![]() such that every positive solution of this system satisfies
such that every positive solution of this system satisfies![]() , otherwise, the system is impermanent.
, otherwise, the system is impermanent.
Lemma 3.2. (See [30] ). If ![]() for all
for all ![]() and if
and if![]() ,
, ![]() then the system
then the system
![]() (3.1)
(3.1)
has a unique positive T-periodic solution which is globally asymptotically stable.
Lemma 3.3. (See [31] ). System
![]() (3.2)
(3.2)
has a unique positive T-periodic solution ![]() which is globally asymptotically stable with respect to
which is globally asymptotically stable with respect to![]() .
.
Lemma 3.4. There exists a positive constant![]() , such that
, such that
![]() (3.3)
(3.3)
for all the solution of system (3.2) with respect to![]() .
.
Proof. Let ![]() be the any solution of system (3.2). By Lemma 3.3, system (3.2) has a unique globally attractive positive T-periodic solution
be the any solution of system (3.2). By Lemma 3.3, system (3.2) has a unique globally attractive positive T-periodic solution![]() . From the global attractivity of
. From the global attractivity of
![]() , for any positive constant
, for any positive constant ![]() (
(![]() ), there exists a
), there exists a![]() , such that for all
, such that for all ![]()
![]() . (3.4)
. (3.4)
By applying (3.4), we obtain![]() ,
, ![]() ,
,![]() .
.
Let
![]() . (3.5)
. (3.5)
We have![]() ,
,![]() .
.
Theorem 3.5. System (2.5) is permanent and has at least one positive T-periodic solution provided
![]() , (3.6)
, (3.6)
where
![]() ,
, ![]() ,
,
where ![]() is the unique positive periodic solution of system (3.2) given by Lemma 3.3 and
is the unique positive periodic solution of system (3.2) given by Lemma 3.3 and ![]() is the upper bound of system (3.2) given by Lemma 3.4 and defined by equation (3.5).
is the upper bound of system (3.2) given by Lemma 3.4 and defined by equation (3.5).
We need the following propositions to prove Theorem 3.5.
Proposition 3.6. For all the solutions of system (2.5) with initial conditions (2.3) and (2.4), we have
![]() , where
, where ![]() is the upper bound of system (3.2) given by Lemma 3.4 and defined by equation (3.5). Furthermore, there exists a positive constant
is the upper bound of system (3.2) given by Lemma 3.4 and defined by equation (3.5). Furthermore, there exists a positive constant![]() , such that
, such that![]() .
.
Proof. Obviously, ![]() is a positively invariant set of system (2.5). Given any solution
is a positively invariant set of system (2.5). Given any solution ![]() of system (2.5) with initial conditions (2.3) and (2.4), we have
of system (2.5) with initial conditions (2.3) and (2.4), we have
![]() ,
,
![]() .
.
Consider the following auxiliary system
![]() (3.7)
(3.7)
By Lemma 3.3, system (3.7) has a unique globally attractive positive T-periodic solution![]() . Let
. Let ![]() be the solution of system (3.7) with
be the solution of system (3.7) with![]() . By the vector comparison theorem [32] , we have
. By the vector comparison theorem [32] , we have
![]() (3.8)
(3.8)
By applying (3.8) and Lemma 3.4, we obtain![]() ,
,![]() .
.
In addition, from the third equation of (2.5) we have
![]() .
.
Consider the following auxiliary equation:
![]() . (3.9)
. (3.9)
According to the condition (3.6), we have
![]() . (3.10)
. (3.10)
By (3.10) and Lemma (3.2), we obtain that system (3.9) has a unique positive T-periodic solution ![]() which is globally asymptotically stable. Then, for the above
which is globally asymptotically stable. Then, for the above ![]() given in (3.4), there exists a
given in (3.4), there exists a![]() , such that
, such that
![]() (3.11)
(3.11)
By applying (3.11), we obtain![]() ,
,![]() .
.
Set![]() , then
, then
![]() . (3.12)
. (3.12)
This completes the proof of Proposition 3.6. □
Proposition 3.7. There exists a positive constant![]() , such that
, such that![]() ,
,![]() .
.
Proof. By Proposition 3.6, there exists a positive ![]() such that
such that![]() ,
,![]() . Hence, from the first and second equations of system (2.5), we obtain
. Hence, from the first and second equations of system (2.5), we obtain
![]() ,
,
![]() ,
,
for![]() . By Lemma 3.3, the following auxiliary system
. By Lemma 3.3, the following auxiliary system
![]() (3.13)
(3.13)
has a unique global attractive positive T-periodic solution![]() . Let
. Let ![]() be the solution of system (3.13) with
be the solution of system (3.13) with![]() , by the vector comparison theorem [32] , we obtain
, by the vector comparison theorem [32] , we obtain
![]() (3.14)
(3.14)
Moreover, from the global attractivity of![]() , there exists a
, there exists a![]() , such that
, such that
![]() ,
,![]() . (3.15)
. (3.15)
Combined (3.14) with (3.15), we have
![]() ,
,![]() (3.16)
(3.16)
Therefore, ![]() ,
,![]() . This completes the proof of Proposition 3.7. □
. This completes the proof of Proposition 3.7. □
Proposition 3.8. Suppose that (3.6) holds, then there exists a positive constant![]() , such that any solution
, such that any solution ![]() of system (2.5) with initial conditions (2.3) and (2.4) satisfies
of system (2.5) with initial conditions (2.3) and (2.4) satisfies
![]() . (3.17)
. (3.17)
Proof. By assumption (3.6), we can choose arbitrarily small constant ![]() (without loss generality, we
(without loss generality, we
assume that![]() , where
, where ![]() is the unique positive periodic solution of
is the unique positive periodic solution of
system (3.2)), such that
![]() , (3.18)
, (3.18)
where
![]()
![]() ,
,![]() .
.
Consider the following system with a parameter![]() ,
,
![]() ,
,
![]() , (3.19)
, (3.19)
By Lemma 3.3, system (3.19) has a unique positive T-periodic solution![]() , which is globally
, which is globally
attractive. Let ![]() be the solution (3.19) with initial condition
be the solution (3.19) with initial condition![]() .
.
Then, for the above![]() , there exists a sufficiently large
, there exists a sufficiently large ![]() such that
such that![]() , for
, for![]() ,
,
![]() .
.
Using the continuity of the solution in the parameter, we have ![]() uniformly in
uniformly in ![]() as
as
![]() . Hence, there exists a
. Hence, there exists a ![]() such that
such that![]() , for
, for![]() ,
, ![]() ,
,
![]() .
.
So, we get![]() . Choosing a constant
. Choosing a constant ![]() (
(![]() ,
,
![]() ), we obtain
), we obtain
![]() ,
, ![]() ,
,![]() . (3.20)
. (3.20)
Suppose that the conclusion (3.17) is not true, then there exists ![]() such that
such that
![]() ,
,
where ![]() is the solution of system (2.5) with initial condition
is the solution of system (2.5) with initial condition
![]() ,
,![]() . So, there exists
. So, there exists ![]() such that
such that
![]() ,
,![]() . (3.21)
. (3.21)
By applying (3.21), from the first and second equation of system (2.5), we have
![]() ,
,
![]() ,
,
for all![]() . Let
. Let ![]() be the solution of system (3.19) with
be the solution of system (3.19) with ![]() and
and
![]() , then we have
, then we have![]() ,
, ![]() ,
,![]() . By the global
. By the global
asymptotic stability of![]() , for the given
, for the given![]() , there exists
, there exists![]() , such that
, such that
![]() ,
, ![]() ,
,![]() .
.
So,
![]() ,
, ![]() ,
,![]() .
.
By using (3.20), we obtain
![]() ,
, ![]() ,
,![]() . (3.22)
. (3.22)
Therefore, by using (3.21) and (3.22), for ![]() it follows
it follows
![]() (3.23)
(3.23)
Integrating (3.23) from ![]() to
to ![]() yields
yields
![]()
Thus, from (3.18) we know that ![]() as
as![]() , which is a contradiction. The proof is com- plete. □
, which is a contradiction. The proof is com- plete. □
Proposition 3.9. Suppose that (3.6) holds, then there exists a positive constant![]() , such that any solution
, such that any solution ![]() of system (2.5) with initial conditions (2.3) and (2.4) satisfies
of system (2.5) with initial conditions (2.3) and (2.4) satisfies
![]() . (3.24)
. (3.24)
Proof. Suppose that (3.24) is not true, then there exists a sequence![]() , such that
, such that
![]() ,
, ![]()
On the other hand, by Proposition 3.8, we have
![]() ,
, ![]()
Hence, there exist time sequences ![]() and
and ![]() satisfying
satisfying
![]() ,
, ![]() ,
, ![]() as
as![]() ,
,
and
![]() ,
, ![]() ,
, ![]() ,
,![]() . (3.25)
. (3.25)
By Proposition 3.6, for a given positive integer m, there exist a![]() , such that for all
, such that for all ![]()
![]() ,
,![]() .
.
Because of ![]() as
as![]() , there is a positive integer
, there is a positive integer![]() , such that
, such that ![]() as
as![]() , hence
, hence
![]() , (3.26)
, (3.26)
for![]() ,
,![]() . Integrating (3.26) from
. Integrating (3.26) from ![]() to
to ![]() yields
yields
![]() ,
,
or
![]() ,
,![]() .
.
Thus, from the boundedness of![]() , we have
, we have
![]() as
as![]() ,
,![]() . (3.27)
. (3.27)
By (3.18) and (3.27), there exist constants ![]() and
and![]() , such that
, such that
![]() ,
, ![]() , (3.28)
, (3.28)
and ![]() for
for![]() ,
, ![]() and
and![]() . (3.28) implies that
. (3.28) implies that
![]() ,
, ![]() , (3.29)
, (3.29)
for![]() ,
,![]() . In addition, for
. In addition, for![]() , we have
, we have
![]() ,
,
![]() .
.
Let ![]() be the solution of (3.19) with
be the solution of (3.19) with ![]() and
and![]() , then by applying
, then by applying
comparison theorem, we have
![]() ,
,![]() .
.
By using Propositions 3.6 and 3.7, there exists a large enough ![]() such that
such that
![]() ,
,
for![]() . For
. For![]() , system (3.19) has a unique positive T-periodic solution
, system (3.19) has a unique positive T-periodic solution ![]() which is global stability. According to the periodicity of (3.19), we have the periodic solution
which is global stability. According to the periodicity of (3.19), we have the periodic solution ![]() is uniformly asymptotically stable in the compact set
is uniformly asymptotically stable in the compact set![]() . Hence, for the given
. Hence, for the given ![]() in Proposition 3.8, there exists
in Proposition 3.8, there exists![]() , which is independent of m and q, such that
, which is independent of m and q, such that
![]() ,
,![]() .
.
Therefore, by using (3.20),
![]() ,
,![]() .
.
According to (3.27), there exists a positive integer ![]() such that
such that ![]() for
for ![]() and
and![]() . Thus, we obtain
. Thus, we obtain
![]() ,
, ![]() , (3.30)
, (3.30)
as ![]() and
and![]() . Therefore, by using (3.29) and (3.30), from the third equation of system (2.5), we have
. Therefore, by using (3.29) and (3.30), from the third equation of system (2.5), we have
![]() ,
,![]() . (3.31)
. (3.31)
Integrating (3.31) from ![]() to
to ![]() leads to
leads to
![]() ,
,
that is,
![]() .
.
This is a contradiction. This completes the proof of Proposition 3.9. □
Proof of Theorem 3.5. By using Propositions 3.6-3.9, system (2.5) is permanent. Using result given by Teng and Chen in [33] , we obtain system (2.5) has at least one positive T-periodic solution. This completes the proof of Theorem 3.5.
4. Extinction
In this section, we investigate the extinction of the predator population in system (2.5) with initial conditions (2.3) and (2.4) under some condition.
Theorem 4.1. Suppose that
![]() . (4.1)
. (4.1)
where
![]() ,
, ![]() ,
,
where ![]() is the unique positive periodic solution of system (3.2) given by Lemma 3.3 and
is the unique positive periodic solution of system (3.2) given by Lemma 3.3 and ![]() is the lower bound of system (3.2) given by Proposition 3.7 and defined by equation (3.16), then
is the lower bound of system (3.2) given by Proposition 3.7 and defined by equation (3.16), then![]() .
.
Proof. According to (4.1), for every given positive constant ![]()
![]() , there exists
, there exists ![]()
![]() and
and ![]() such that
such that
![]() (4.2)
(4.2)
where
![]() ,
,![]() .
.
From the first and second equations of system (2.5), we have
![]() ,
,
![]() .
.
Hence, for the above ![]() there are exists a
there are exists a![]() , such that
, such that
![]() ,
, ![]() ,
,![]() . (4.3)
. (4.3)
It follows from (4.2) and (4.3) that for![]() ,
,
![]() . (4.4)
. (4.4)
Firstly, we show that exists a![]() , such that
, such that![]() . Otherwise, by (4.4), we have
. Otherwise, by (4.4), we have
![]()
That is to say![]() . This is a contradiction.
. This is a contradiction.
Secondly, we show that
![]() , for
, for![]() , (4.5)
, (4.5)
where
![]() ,
,
is bounded for![]() . Otherwise, there exists a
. Otherwise, there exists a ![]() such that
such that![]() .
.
By the continuity of![]() , there must exists a
, there must exists a ![]() such that
such that ![]() and
and ![]() for
for
![]() . Let
. Let ![]() be the nonnegative integer such that
be the nonnegative integer such that![]() . According to
. According to
(4.3), we have
![]()
which is a contradiction. This shows that (4.5) holds. By the arbitrariness of![]() , it immediately follows that
, it immediately follows that ![]() as
as![]() . This completes the proof of Theorem 4.1. □
. This completes the proof of Theorem 4.1. □
5. Examples
In this section, we give some examples to illustrate the feasibility of our main results in Theorems 3.5 and 4.1.
Example 5.1. Let
![]() (5.1)
(5.1)
In this case, system (3.2) given by Lemma 3.3 has a unique positive periodic solution
![]() . Let
. Let![]() , it is easy to know
, it is easy to know![]() . By a simple
. By a simple
calculation, we have
![]() .
.
According to Theorem 3.5, system (2.5) with the above coefficients is permanent and admits at least one positive 2p-periodic solution for any nonnegative 2p-periodic function![]() . Figure 1 shows the dynamic behavior of system (2.5) with the above coefficients and
. Figure 1 shows the dynamic behavior of system (2.5) with the above coefficients and![]() . Figure 2 shows the dynamic behavior of system
. Figure 2 shows the dynamic behavior of system
(2.5) with the above coefficients and![]() . From Theorem 3.5, we know that the overcrowding rate
. From Theorem 3.5, we know that the overcrowding rate ![]() of the predator population has no influence on the permanence of system (2.5). However, from Figure 1 and Figure 2, we can know that the overcrowding rate influence the density of the predator population. Figure 1 demonstrates that the predator species is at low density with the overcrowding effect; Figure 2 shows that the predator population is at high density without the overcrowding effect. According to the biological viewpoint, the overcrowding effect restricts population growth.
of the predator population has no influence on the permanence of system (2.5). However, from Figure 1 and Figure 2, we can know that the overcrowding rate influence the density of the predator population. Figure 1 demonstrates that the predator species is at low density with the overcrowding effect; Figure 2 shows that the predator population is at high density without the overcrowding effect. According to the biological viewpoint, the overcrowding effect restricts population growth.
Example 5.2. In system (2.5), let![]() ,
,![]() . And the other parameters are given by equations (5.1). In the same way as Example 5.1, it is easy to verify that system (2.5) is permanent at this case. The dynamic behavior of system (2.5) in such conditions is given by Figure 3. Compared with Figure 1, we can know that the predator is at high population density if the predator prefers the immature prey to the mature, whereas it is at lower population density. According to the biological viewpoint, prey vulnerability is a major factor influencing the predator preference [34] . Compared with the mature prey, the immature is more easily captured by the predator. Then, the predator population density is high if the predator prefers the immature prey.
. And the other parameters are given by equations (5.1). In the same way as Example 5.1, it is easy to verify that system (2.5) is permanent at this case. The dynamic behavior of system (2.5) in such conditions is given by Figure 3. Compared with Figure 1, we can know that the predator is at high population density if the predator prefers the immature prey to the mature, whereas it is at lower population density. According to the biological viewpoint, prey vulnerability is a major factor influencing the predator preference [34] . Compared with the mature prey, the immature is more easily captured by the predator. Then, the predator population density is high if the predator prefers the immature prey.
Example 5.3. In system (2.5), let![]() ,
,![]() . And the other parameters are given by equations (5.1). In this case, by a simple calculation, we obtain
. And the other parameters are given by equations (5.1). In this case, by a simple calculation, we obtain
![]() .
.
According to Theorem 4.1, system (2.5) is impermanent and the predator population is extinction. Numerical simulation given in Figure 4 also confirms the result.
6. Conclusions
In this paper, we propose and analyze a periodic predator-prey system with time delay and prey stage-structured preference by the predator. The permanence and existence of positive periodic solutions of system (2.5) are explored. The conditions for the impermanence of the system and the extinction of the predator population are obtained. By Lemma 3.3, we know that ![]() is the globally attractive periodic solution of system (2.5) without predation. Hence, the condition (3.6) implies that system (2.5) is permanence if the death rate of the predator population is small enough. Numerical simulations not only show the consistency with the theoretical analysis but also exhibit other interesting biological phenomenon. From Example 5.1, we know that the predator population is at high density without the overcrowding effect. By Examples 5.1 and 5.2, we get that the predator’s preference to the immature prey is beneficial for itself development. This is because the immature prey is
is the globally attractive periodic solution of system (2.5) without predation. Hence, the condition (3.6) implies that system (2.5) is permanence if the death rate of the predator population is small enough. Numerical simulations not only show the consistency with the theoretical analysis but also exhibit other interesting biological phenomenon. From Example 5.1, we know that the predator population is at high density without the overcrowding effect. By Examples 5.1 and 5.2, we get that the predator’s preference to the immature prey is beneficial for itself development. This is because the immature prey is
more palatable and more easily captured by the predator than the mature. Example 5.3 illustrates the correctness of Theorem 4.1.
We would like to mention here that we are unable to solve the following questions:
1) How many positive periodic solutions exist in system (2.5)?
2) Is the solution global attractivity if system (2.5) has only one positive periodic solution?
We leave these for future work.
Acknowledgements
The authors express sincere gratitude to the anonymous referees for their helpful comments and suggestions that led to an improvement of our original manuscript.
Funding
This work was supported by the Major Project of Sichuan University of Arts and Science (Grant No. 2014Z005Z), by the General Project of Educational Commission in Sichuan Province (Grant No. 16ZB0357).
NOTES
![]()
*Corresponding author.