American Journal of Molecular Biology
Vol.07 No.01(2017), Article ID:72869,10 pages
10.4236/ajmb.2017.71001
Studies on Trans-Generational Transcriptional Silencing of cry1Ac Gene in Tobacco Transgenics
Madhurima Kahali, Kamlesh Kumar Soni, Pradeep Kumar Burma*
Department of Genetics, University of Delhi South Campus, New Delhi, India

Copyright © 2017 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: November 19, 2016; Accepted: December 17, 2016; Published: December 20, 2016
ABSTRACT
Developing transgenics that express high levels of Cry1Ac protein, and at the same time, are phenotypically normal, has not been an easy task to achieve. It has been routinely observed that most of the transgenic plants that survive, show no or extremely low levels of Cry1Ac protein. However, all of these plants do express the selectable marker, nptII gene. In the present study, we record an interesting observation of how one of the genes (cry1Ac) on a single T-DNA fragment is selectively silenced, keeping the expression of the other gene (nptII) intact. Further, this silenced state is inherited.
Keywords:
Cry1Ac, Transcriptional Silencing, Bt Crops, Gene Silencing

1. Introduction
Insect resistant Bt cotton is the only commercialized genetically modified crop in India so far. From a few hectares when introduced in 2002, the transgenic cotton now occupies about 95% of the total cotton-growing acreage of 12.25 million hectares [1] in India. Cotton is an economically important crop for India. The crop shows high susceptibility to lepidopterans, the major one in India being Helicoverpa armigera. The use of Bt cotton, which specifically kills these lepidopteran pests, has been a boon to the cotton industry. The insecticidal transgenic crop, utilizes the δ-endotoxin produced by the ubiquitously present, Gram-positive bacteria, Bacillus thuringiensis to kill the attacking pests. Bacillus thuringiensis contains an array of insecticidal toxins including Cry proteins, cytolysins (Cyt toxins), enterotoxins, chitinases, hemolysins, phospholipases etc. [2] . Bt cotton exploits the insecticidal activity of the Cry proteins. This is a very robust technique as the cry genes are extremely specific in terms of their target insect group.
Following the success of Bt cotton, many laboratories ventured into developing new Bt lines with high toxin levels. However, this hasn’t been an easy feat to achieve, and so far very few Bt events have been commercialized-though, hybrids made from these events are quite many. Many laboratories have reported difficulties in developing transgenics with cry genes [3] [4] [5] .
It has been shown in a study by our laboratory that the expression of cry1Ac gene has adverse effects on the in vitro regeneration and growth of transgenic plants, in studies involving cotton and tobacco [6] . During the experiments undertaken in cotton, a strong correlation was observed between level of expression of Cry1Ac protein and phenotypic abnormality, meaning thereby, that the transgenic plants exhibiting high levels of the Cry1Ac protein showed abnormalities such as stunted growth, no boll formation, lack of seed setting etc. In this work [6] , an important observation was that, out of all the T0 independent transgenic lines developed in cotton, with the binary vector represented in Figure 1, only 15% of the lines were observed to show detectable levels of Cry1Ac protein as tested by ELISA, though all of these lines expressed the marker gene nptII. This analysis was carried out in T0 transgenics. Thus, in these transgenic lines, the expression of the cry1Ac gene was silenced, while the neighbouring selection marker, nptII seemed to be active.
Further, Southern hybridization showed that in about ~29% of the transgenics analysed, the cry1Ac cassette was physically deleted, although the nptII cassette was found to be present [6] . Such observation had not been reported earlier in studies on developing transgenic lines with different genes. Interestingly, in these transgenic lines the cry1Ac cassette was placed towards the right border (RB) of the T-DNA, while the selection marker (nptII) was placed towards the left border (LB). It is well established that T-DNA transfer proceeds from RB to LB, and although incomplete transfers can take place at both the borders, the placement of the marker at the LB ensures the presence of the passenger gene in most of the cases [7] .
The transformation experiments, using the same construct as used in cotton (Figure 1), when repeated in tobacco (Nicotianata bacum cv. Xanthi) showed similar results. Again, a major observation was that a large percentage of the developed kanamycin resistant transgenic tobacco did not show observable level of Cry1Ac protein.
In summary, in the previous work there were two different kinds of kanamycin resistant (KanR) transgenic tobacco lines observed, ones that expressed cry1Ac gene (hereafter called as expressors), and those that did not express cry1Ac (non-expressors). The present study was undertaken to test the inheritance of the expression status in these lines in the T1 generation, as well as to try and identify the possible reasons for the interesting observation of selective silencing of the cry1Ac gene, keeping the selection marker (nptII) active. For the present study progeny of 6 independent tobacco T0
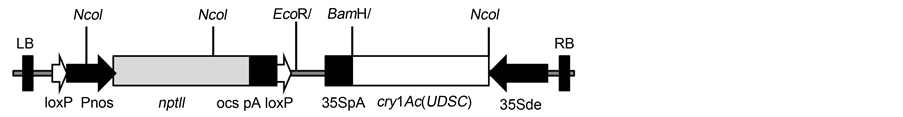
Figure 1. A vector map of the construct used for developing transgenics in tobacco and cotton, used by Rawat et al. 2011. The construct carries the cry1Ac (UDSC) gene driven by a CaMV 35S double enhancer promoter at the right border, and a selection marker nptII gene driven by the nos promoter at the left border.
transgenic lines were selected for further analysis viz., three expressor lines viz. E1.7, E2.19 and E3.39, and three non-expressor lines viz. NE1.59, NE1.35 and NE1.08 (from the previous study [6] ). All the six transgenic lines chosen were developed through Agrobacterium-mediated transformation of tobacco, using the construct described in Figure 1. The expressor lines, E1.7, E2.19 and E3.39, expressed both cry1Ac and nptII genes. The levels of Cry1Ac protein expressed in these lines were, ~0.48 ppb/µg total protein for E1.7, ~1.3 ppb/µg total protein for E2.19, and, ~3.43 ppb/µg total protein for E3.39, respectively. The non-expressor lines, NE1.59, NE1.35 and NE1.08 showed no detectable levels of Cry1Ac protein though they were resistant to kanamycin.
2. Materials and Methods
2.1. Segregation Analysis of T1 Generation Seeds
Six independent tobacco transgenic lines were selected, three expressor lines viz. E1.7, E2.19 and E3.39, and three non-expressor lines viz. NE1.59, NE1.35 and NE1.08 (from the previous study [6] ). T1 seeds of these lines were germinated on MS (Murashige and Skoog) media [8] , at pH 5.9 - 6.0 with kanamycin (100 mg/L) as selection agent, to study the segregation pattern of the transgene. Seeds that germinated on kanamycin was scored as resistant (KanR) and those that did not as sensitive (KanS) Segregation analysis was done using the χ2 test, where the observed values were compared to the expected values, corresponding to the integration of the transgene at a single locus following a 3:1 Mendelian ratio, of KanR:KanS seedlings. Progeny of lines with integration of transgene at single locus were taken for further analysis.
2.2. Analysis of Presence/Absence of Cry1Ac Protein in Transgenics by Qualitative ELISA
The expression of the Cry1Ac protein was studied in the T1 progeny lines by Qualitative ELISA. Twelve randomly selected KanR progeny lines of NE1.59, NE1.35. E1.7, E2.19 and E3.39 were selected for this analysis. Analysis of Cry protein was carried out on the first fully expanded leaf of the putative transgenic. This was carried out using Sandwich Enzyme Linked Immunosorbent Assay (ELISA) from Envirologix, USA. This was carried out using Cry1Ac/Ab ELISA kit, Amar Immunodiagnostics, India.
2.3. Analysis of Cry1Ac Protein Level in Different Progeny Lines by Quantitative ELISA
The progeny lines of the three Cry1Ac expressing lines, i.e. E2.19, E1.7 and E3.39 ELISA were analyzed for their protein levels by Quantitative ELISA kit from Envirologix, USA. Estimation of Cry1Ac protein was carried out using 0.5 µg and 1.0 µg of total protein. The total protein concentration in each sample was determined by Bradford method [9] , using known concentrations of BSA as standard.
2.4. Analysis of Presence/Absence of cry1Ac and nptII Gene Cassettes Using PCR
Six progeny lines of NE1.35 were randomly selected and PCR amplifications were carried out on the genomic DNA from different regions, these regions included, cry1Ac complete cassette, cry1Ac 5’ end, cry1Ac 3’ end, nptII gene and the 35S de promoter region. This was done to make sure that there was no truncation or deletion in the Cry1Ac protein, which resulted in the silencing. All the amplicons were sequenced to check for the presence of any random mutation. The sequences of the oligonucleotides used for amplification is presented in Table 1.
2.5. Analysis of Transcript Level by RT-PCR
Total RNA was extracted using “Spectrum Plant RNA isolation kit” from Sigma, following the manufacturer’s instructions. cDNA was synthesized using QuantiTect Reverse Transcription kit (Qiagen). The cDNA synthesis was carried out in a reaction volume of 20 µl.
Amplifications were done on the selectable marker nptII gene, along with both 3’ and 5’ends of the cry1Ac gene to make sure that there was no truncation, using gene specific primers. Ubiquitin-conjugating enzyme (UCE) specific amplifications were carried out as internal control. The sequences of oligonucleotides that have been used for the amplifications are the same as used for the PCR amplifications on genomic DNA (Table 1).
3. Results
3.1. Segregation Analysis of the T1 Generation Seeds
Based on the χ2 test, it was observed that 5 of the 6 lines had integration of the transgene at a single locus. The results have been summarized in Table 2.
3.2. Analysis of Presence/Absence of Cry1Ac Protein in T1 by Qualitative ELISA
Approximately 12 T1 progeny lines of the independent T0 lines were analysed for the

Table 1. Sequences of primers and their TM used for amplification of different fragments of the integrated transgenes. The expected size of amplicons is also shown.
Table 2. Segregation analysis of T1 progeny lines of 6 independent transgenic lines.
*χ2 fits the 3:1 ratio.
presence of Cry1Ac protein. 0.5 μg and 1.0 µg of total soluble protein were used for the analysis. It was observed that all progeny of the non-expressors showed absence of the Cry1Ac protein, while all the progeny of the expressors, except for two lines of E2.19, showed the presence of Cry1Ac protein.
3.3. Analysis of Cry1Ac Protein Level by Quantitative ELISA in T1 Progeny of Expressors
To analyse the level of Cry1Ac protein in T1 progeny of expressor lines, quantitative ELISA using 0.5 μg and 1.0 μg of total soluble protein was carried out (Figure 2). It was observed that in case of E1.7, only one out of the twelve progeny analyzed, showed detectable but very low level of Cry1Ac protein. This line was a low expressor (0.48 ppb/µg) even in T0 generation, and the Cry1Ac locus seems to have been silenced in the T1 generation. However, all these lines rooted on kanamycin 100 mg/L (selection), indicating their transgenic nature. For line E2.19, out of the twelve progeny lines analyzed, seven showed detectable levels of Cry1Ac protein, the expression levels ranging from 0.022 to 0.955 ppb/µg of total protein. The parent line had moderate expression levels of Cry1Ac of ~1.3 ppb/µg. In the case of line E3.39, the parental line was a high expressor of Cry1Ac (3.43 ppb/µg), all the progeny lines showed detectable levels of Cry1Ac expression, the expression levels ranged from 0.181 to 6.839 ppb/µg of total protein.
3.4. Analysis of Presence/Absence of cry1Ac and nptII Gene Cassettes in T1 Progeny of Non-Expressor Lines
Six T1 progeny of the non-expressor line 1.35 were analysed for the presence or absence of the transgene cassette by PCR. Primers designed from different regions of the cry1Ac (i.e promoter, 5’ and 3’ halves of the cry1Ac gene) and nptII gene cassettes (Table 1) were used to study the structural integrity of the transgenes. It was observed that all the cassettes were present and there was no aberration in the integrated cassette (Figure 3). This observation was different from a previous observation made in cotton [6] , were a large number of transgenics were observed not to carry the cry1Ac gene while retaining

Figure 2. Levels of Cry1Ac protein as detected by Quantitative ELISA in the T1 progeny of different expressor lines.
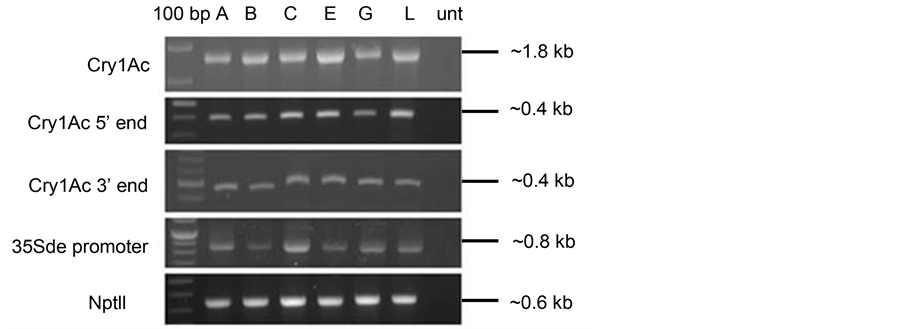
Figure 3. PCR amplifications with-cry1Ac gene (full length), 5’ end of cry1Ac (1 - 401 bp), 3’ end of cry1Ac (1381 - 1848 bp), 35S double enhancer promoter and nptII specific primers, on genomic DNA of 6 selected progeny lines of non-expressor NE1.35 viz. 1.35A, 1.35B, 1.35C, 1.39E, 1.35G and 1.35L, along with untransformed tobacco as control. Lane 1 is 100 bp ladder (New England Biolab, USA). After an initial denaturation at 94˚C for 5 minutes, 30 cycles of denaturation at 94˚C for 30 s, annealing at temperatures according to Tm of primer pairs for 30 s and extension at 72˚C for 30 s was carried out. This was followed by a final extension for 7 min at 72˚C.
the nptII gene.
The amplified fragments of the 35S double enhancer promoter regions, as well as the cry1Ac gene cassette were sequenced for any possible mutations (small deletions) in the promoter or possible nonsense mutations in the coding frame, which could account for non-expression of the Cry1Ac protein in these lines. However, no such aberration or mutation was observed.
3.5. Analysis of Transcript Level by Semi-Quantitative RT-PCR
Six T1 progeny each of an expressor line (E3.39) and a non-expressor line (NE1.35) were analysed for the presence or absence of the transcripts encoded by the transgenes, by RT-PCR (Figure 4). It was observed that all the analysed lines showed the presence of the nptII transcript. Progeny of the E3.39 line showed the presence of cry1Ac transcript. However, the T1 progeny of line NE1.35 mostly did not show the presence of cry1Ac, while all these lines showed appreciable levels of the nptII transcript. One of the lines (NE1.35B) showed amplification at the 5’ end, but none using the primers towards the 3’ end. The observations were reproducible with biological replicates.
4. Discussion
One of the major impediments in developing insect resistant transgenics with cry1Ac gene is to obtain a line with high levels of Cry1Ac protein, without the phenotype of the transgenic line being adversely affected [6] . It has been hypothesised that due to these adverse effects there is an enrichment of transgenic events which either do not express or express extremely low levels of the Cry1Ac protein. Thus, a large proportion of the developed transgenic lines are observed to have silenced the cry1Ac gene. The present work, a continuation of a previous work from the laboratory [6] , analyzes the status of the expression of cry1Ac gene in the T1 generation of transgenic tobacco plants and tries to identify the reason for the same.
The present study showed that transcriptional silencing of the cry1Ac gene was the reason for absence of the protein in the non-expressors. The structural integrity of the gene was found to be intact in these non-expressor lines. The non-expressors were found to lack the transcripts. Interestingly, the selectable marker, NptII, which was a part of the same T-DNA fragment, was not silenced in these lines. Thus, this is an
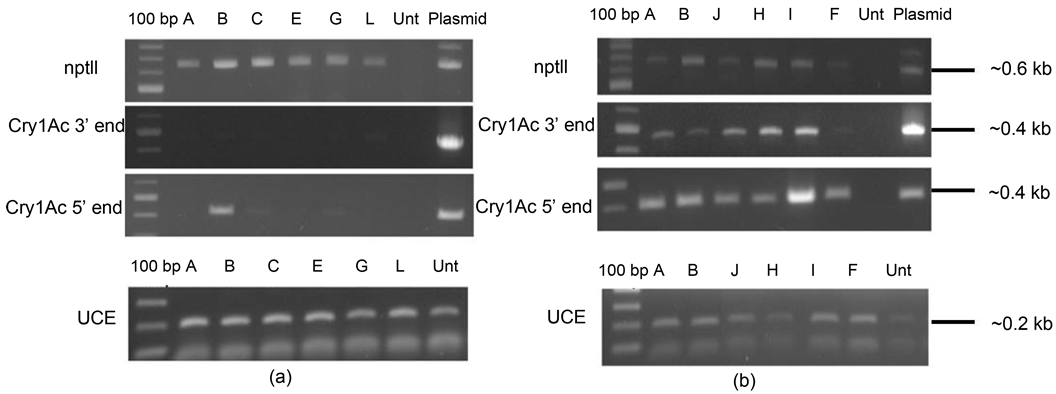
Figure 4. RT-PCR amplifications with-nptII, cry1Ac 3’ end (1 - 401 bp), cry1Ac 5’ end (1381 - 1848 bp) and ubiquitin-conjugate enzyme specific primers, on the cDNA of: (a) 6 selected progeny lines of non-expressor NE 1.35 viz. 1.35A, 1.35B, 1.35C, 1.39E, 1.35G and 1.35 L, along with untransformed tobacco and plasmid DNA as control, (b) 6 selected progeny lines of E3.39 viz. 3.39A, 3.39B, 3.39J, 3.39H, 3.39I and 3.39F, along with untransformed tobacco and plasmid DNA as control. After an initial denaturation at 94˚C for 5 minutes; 25, 30 and 35 cycles respectively, of denaturation at 94˚C for 30 s; annealing at temperatures according to Tm of primer pairs for 30 s and extension at 72˚C for 30 s was carried out. This was followed by a final extension for 7 min at 72˚C.
example of transcriptional silencing of one specific gene in a transgene construct.
Stability of transgene is a big challenge that plant biologists have to grapple with. The phenomenon of transgene silencing in higher plants is widely known. It is also known that gene silencing in plants is a way of warding off potential threat-much like the immune response in animals. Transgenes that resemble cellular invaders are acted upon by the host defense system, and silenced [10] . There are two well-known mechanisms through which silencing takes place. The first is associated with de novo DNA methylation, while, the second is post-transcriptional silencing through RNA degradation [11] . Apart from the above two, there can also be mutations in the transgene that result in silencing. Truncation in the T-DNA integration is one of the most likely causes of transgene inactivity [12] . As far as the present study is concerned, no such truncations have been observed. Neither have we observed any point mutations, as analysed by de novo DNA sequencing. So, we could rule out this mechanism of silencing in the plants under study.
The present study also showed that the silencing of the cry1Ac gene is heritable. The T1 progeny lines of the non-expressor lines, continued to not express the cry1Ac gene, while expressing the linked selectable marker, nptII. Further, the silencing of the cry1Ac gene seems to be at the transcriptional level. All these observations indicate an epigenetic silencing of Cry1Ac. As discussed earlier, transgene silencing is a defense mechanism for plants. A number of studies have shown that plants silence potentially hazardous genes, or genes that bring about stress to the plant, in order to ensure their healthy development. This kind of epigenetic silencing is also passed on across generations. What we observe in our study, could also be a similar case. Studies have shown that level of expression among plants transformed with the same construct can differ considerably, and in many cases this has nothing to do with copy number [13] [14] . Nonetheless, nothing has been discussed about such variation, as observed in our study, with regard to the expression of transgenes on the same T-DNA fragment. Though a lot has been discussed in literature about transgene silencing, the phenomenon of silencing of one of the transgenes on a single T-DNA fragment, has not been talked about. It would be interesting to delve deeper into this area. It is also a noteworthy observation, that the plants that had the cry1Ac gene silenced―were all healthy. This is in contrast to the plants that expressed high levels of Cry1Ac protein―most of these plants show phenotypic abnormalities. A study carried out by Barton et al. 1987 [3] demonstrated that expression of intact Cry1Ac protein in tobacco was lethal to the plant cells. The only plants that regenerated were the ones that either contained a truncated or an inactive Cry1Ac protein. A similar observation was made by Rawat et al. 2011 [6] , where it was seen that transgenic tobacco plants with no or low Cry1Ac levels would show no apparent abnormalities, while the ones that expressed high levels of Cry1Ac would either not grow at all, or, show both vegetative and reproductive abnormalities. These observations further point that the selective silencing of the cry1Ac gene might have been the plant’s way to combat stress caused by the Cry1Ac toxin. It could also be argued that silencing can occur randomly. In this specific example the T-DNA contains two expression cassettes viz., cry1Ac and nptII. During transformation of tobacco with this construct, there could be events where both the cassettes are silenced and also those where only one of them is silenced. During the process of transformation and selection, resistance to kanamycin (encoded by nptII gene) acts as a positive selection; thus all selected transgenic events are expected to show the expression of nptII. On the other hand, presence of Cry1Ac protein adversely affects the growth of the transformants and thus leads to selection of events where the cry1Ac gene is silenced. Though this is an interesting observation, additional experiments need to be carried out, to understand the whole process of the selective silencing better.
Acknowledgements
This work was supported by a grant from Delhi University. MK was supported by a research fellowship from Council of Scientific and Industrial Research (CSIR), Delhi, India.
Cite this paper
Kahali, M., Soni, K.K. and Burma, P.K. (2017) Studies on Trans-Generational Transcriptional Silenc- ing of cry1Ac Gene in Tobacco Transgenics. American Journal of Molecular Biology, 7, 1-10. http://dx.doi.org/10.4236/ajmb.2017.71001
References
- 1. James, C. (2014) Global Status of Commercialized Biotech/GM Crops. ISAAA Brief, 49.
- 2. de Maagd, R.A., Bravo, A. and Crickmore, N. (2001) How Bacillus thuringiensis Has Evolved Specific Toxins to Colonize the Insect World. Trends in Genetics, 17, 193-199.
https://doi.org/10.1016/S0168-9525(01)02237-5 - 3. Barton, K.A., Whiteley, H.R. and Yang, N.S. (1987) Bacillus thuringiensis § Endotoxin Expressed in Transgenic Nicotiana Tabacum Provides Resistance to Lepidopteran Insects. Plant Physiology, 85, 1103-1109.
https://doi.org/10.1104/pp.85.4.1103 - 4. Chakrabarti, S.K., Lutz, K.A., Lertwiriyawong, B., Svab, Z. and Maliga, P. (2006) Expression of the cry9Aa2 Bt Gene in Tobacco Chloroplasts Confers Resistance to Potato Tuber Moth. Transgenic Research, 15, 481-488.
https://doi.org/10.1007/s11248-006-0018-z - 5. Acharjee, S., Sarmah, B.K., Kumar, P.A., Olsen, K., Mahon, R., Moar, W.J., Moore A. and Higgins, T.J.V. (2010) Transgenic Chickpeas (Cicer arietinum L.) Expressing a Sequence-Modified cry2Aa Gene. Plant Science, 178, 333-339.
https://doi.org/10.1016/j.plantsci.2010.02.001 - 6. Rawat, P., Singh, A.K., Ray, K., Chaudhary, B., Kumar, S., Gautam, T., Kanoria, S., Kaur, G., Kumar, P., Pental, D. and Burma, P.K. (2011) Detrimental Effect of Expression of Bt Endotoxin Cry1Ac on in Vitro Regeneration, in Vivo Growth and Development of Tobacco and Cotton Transgenics. Journal of Biosciences, 36, 363-376.
https://doi.org/10.1007/s12038-011-9074-5 - 7. Lee, L.Y. and Gevin, S.B. (2008) T-DNA Binary Vectors and Systems. Plant Physiology, 146, 325-332.
https://doi.org/10.1104/pp.107.113001 - 8. Murashige, T. and Skoog, F. (1962) A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiologia Plantarum, 15, 473-497.
https://doi.org/10.1111/j.1399-3054.1962.tb08052.x - 9. Bradford, M.M. (1976) A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Analytical Biochemistry, 72, 248-254.
https://doi.org/10.1016/0003-2697(76)90527-3 - 10. Park, Y.D., Papp, I., Moscone, E.A., Iglesias, V.A., Vaucheret, H., Matzke, A.J.M. and Matzke, M.A. (1996) Gene Silencing Mediated by Promoter Homology Occurs at the Level of Transcription and Results in Meiotically Heritable Alterations in Methylation and Gene Activity. The Plant Journal, 9, 183-194.
https://doi.org/10.1046/j.1365-313X.1996.09020183.x - 11. Matzke, M.A., Mette, M.F. and Matzke, A.J.M. (2000) Transgene Silencing by the Host Genome Defense: Implications for the Evolution of Epigenetic Control Mechanisms in Plants and Vertebrates. Plant Molecular Biology, 43, 401-415.
https://doi.org/10.1023/A:1006484806925 - 12. Francis, K.E. and Spiker, S. (2005) Identification of Arabidopsisthaliana Transformants without Selection Reveals a High Occurrence of Silenced T-DNA Integrations. The Plant Journal, 41, 464-477.
https://doi.org/10.1111/j.1365-313X.2004.02312.x - 13. Peach, C. and Velten, J. (1991) Transgene Expression Variability (Position Effect) of CAT and GUS Reporter Genes Driven by Linked Divergent T-DNA Promoters. Plant Molecular Biology, 17, 49-60.
https://doi.org/10.1007/BF00036805 - 14. Tizaoui, K. and Kchouk, M.E. (2012) Genetic Approaches for Studying Transgene Inheritance and Genetic Recombination in Three Successive Generations of Transformed Tobacco. Genetics and Molecular Biology, 35, 640-649.
https://doi.org/10.1590/S1415-47572012000400015