American Journal of Plant Sciences
Vol.3 No.6(2012), Article ID:20024,11 pages DOI:10.4236/ajps.2012.36091
A Flower Specific Calcineurin B-Like Molecule (CBL)-Interacting Protein Kinase (CIPK) Homolog in Tomato Cultivar Micro-Tom (Solanum lycopersicum L.)
![]()
Department of Bioresource Science, Faculty of Agriculture, Kyushu University, Fukuoka, Japan.
Email: *yuasa@agr.kyushu-u.ac.jp
Received March 27th, 2012; revised April 10th, 2012; accepted April 30th, 2012
Keywords: Calcium; CBL; CIPK; Flower; SnRK; Tomato
ABSTRACT
Floral and reproductive organs of higher plants are relatively sensitive to biotic and abiotic stresses compared with the vegetative organs. Calcineurin B-like molecule (CBL) interacting protein kinase (CIPK) has appeared to be involved in acquired tolerance and acclimation under environmental stresses such as salinity, drought and chilling. Semi-quantitative RT-PCR using the vegetative and reproductive organs of tomato Micro-Tom (Solanum lycopersicum L.) at the various developmental stages indicated that SlCIPK2 was expressed specifically in the floral organ. An anti-CIPK specific antibody recognized the recombinant SlCIPK2 specifically and cross-reacted to a CIPK-related polypeptide at a significant level in flower, particularly in stamen. The flower specific CIPK was tightly associated with the microsomes. In vitro pull-down assay of the recombinant SlCIPK2 showed that SlCIPK2 interacts with SlCBLs and stress-responsive transcription factors, SlERF7, SlCBF1 and SlAREB1. The present data suggested that the flower-specific CIPK, SlCIPK2, was involved in calcium signaling in tomato via CBLs and stress tolerance possibly mediated by the stressresponsive transcription factors in stamen.
1. Introduction
In higher plants, the floral development is well programmed and plays an important role in plant reproduction. Compared with the vegetative development, the reproductive development is relatively susceptible to environmental stresses such as high temperature, drought and chilling. For an example of tomato races, the fruit production under moderate heat shock is dependent on that genetic background, but there was marginal difference in neither photosynthetic rate nor respiration between the heat tolerant race and the sensitive race, when subjected to high temperature stress. Reduction of tomato fruit yield under high temperature stress was mostly due to failure of pollen development, such as formation of sterile pollen. A chaperone, HSP90, has appeared to be expressed specifically in floral organs of Arabidopsis and rice in particular, in pollen [1,2]. Pollen accumulates osmolites, proline and sucrose at high concentration, indicating that specific protective mechanisms against heat and/or osmotic stress are involved in development of pollen [3].
In plants, complex mechanisms including stress signaling cascades and synthesis of osmolytes have been evolved to survive under various environmental stresses [4]. For molecular and genetic breeding directing to improvement of stress tolerance during reproduction stage of crops, investigation of intracellular mechanisms of stress signaling of floral organs and fruit, which are supposed to be susceptible to various stresses, becomes important in an aspect of agriculture encountering to the situation of global warming.
It is well known that protein phosphorylation/dephosphorylation has an essential role in stress tolerance and adaptation of eukaryotes [5,6]. Recent studies of organ specific SNF1-related protein kinases (SnRKs) revealed that Vicia faba guard cell-specific SnRK2, ABA-activated protein kinase (AAPK) [7], and A. thaliana root tip-specific SnRK2C/SnRK2.8 [8] are involved in closure of guard cells and tolerance of osmotic stress, respectively, and that tomato fruit-specific SnRK2 homolog, SlSnRK2C, are activated by NaCl and chilling [9].
In plant cells, Ca2+ ion is a second messenger involveing the regulation of enzymatic activity and gene expression via calmodulins and Ca2+-dependent protein kinases. Recent studies have clarified that a calcium sensor “Calcineurin B-Like protein” (CBL), containing Ca2+-binding EF-hand motifs, interacts to CBL-Interacting Protein Kinases (CIPKs) [10]. The CIPKs contain a kinase domain which has significant similarity in aminoacid sequence to highly similar to SNF1, at the N-terminus and a specific interacting domain, containing a NAF/FISL motif, associating to CBL at the COOH regulatory domain [10,11]. On the basis of these structural similarities, the CIPKs belongs to the SnRK3 subgroup of plant SNF1-related kinases [12]. Biochemical and genetic research by Arabidopsis mutants have revealed that the CBL-CIPK complex is translocated onto microsomes and implicated in ion homeostasis, solute transport and aquring tolerance under environmental stresses [13,14].
Tomato cultivar Micro-Tom (Solanum lycopersicum L.) is a model crop and suitable for research of fruit, secondary metabolisms and environmental signaling such as pathogenic stress and temperature stresses, etc. We profiled expression levels of CIPK homologs in tomato organs and identified SlCIPK2 as a flower specific CIPK. Immunoblot and pull-down assay indicated that the flowers specific CIPK is localized in stamen, associating to the microsomal fraction and interacts with stress-responsive transcription factors, suggesting involvement of pollen development and protection against environmental stresses.
2. Materials and Methods
2.1. Plant Materials
Seeds of the dwarf tomato (Solanum lycopersicum L. cv. Micro-Tom) were supplied from the Kazusa DNA Research Institute in Japan. Tomato seeds were incubated in 6-cm-square rock wool under continuous light (80 W∙m–2) and fertilized with 1 to 1000-diluted Hyponex (Hyponex Japan, Osaka, Japan). Various organs from 6 - 8 week old tomato plants were frozen in liquid N2 for source of total RNA and protein samples.
2.2. RNA Extraction and Semiquantitative RT-PCR of Tomato CIPK and CBL Orthologs
At least, 9 sequences among “contigs” coding tomato CIPK orthologs were predicted by BLAST search using Arabidopsis CIPK1 [10] and CIPK24/SOS2 [15] as search terms against the Micro-Tom EST database (MiBASE) and Kazusa Tomato Unigene ver. 3 (KTU3) (http://www. kazusa.or.jp/jsol/microtom/indexj.html). The deduced amino acid sequences of the tomato CIPK candidates and Arabidopsis CIPKs (http://mips.gsf.de/proj/thal/db/index.html) were aligned by the CLUSTALW program (http://align. genome.jp/). Eventually, we focused on 4 candidates, SlCIPK1, 2, 8 and SlSOS2, because of putatively higher expression levels of the genes than those of other tomato CIPK orthologs based on numbers of ESTs supporting those contigs in MiBASE. SlCIPK1, SlCIPK2 (KTU3 contig27053, AB674507, this study) and SlCIPK8 were designated corresponding to AtCIPK1, AtCIPK2, AtCIPK8 and AtCIPK24/SOS2, respectively (Figure 1(b)). Similarly, tomato CBL orthologs, SlCBL1, 2, 10 and SlSOS3, were designated corresponding to AtCBL1, AtCBL2, AtCBL10 and AtCBL4/SOS3 (Figure 2(c)).
 (a)
(a) (b)
(b)
Figure 1. The expression profiles of tomato CIPK homologs in various organs of tomato plant. (a) The expression levels of mRNAs of SlCIPK2, SlCIPK1, SlCIPK8, SlSOS2 and SlSOS1 in various organs are shown. Semi-quantitative RT-PCR was carried out at indicated numbers of thermal cycles and with cDNA prepared from various organs and sets of specific primers for detection of tomato CIPK homologs and SlSOS1. SlUbi was used as a standard; (b) A phylogenic tree of CIPKs, SNF1-related kinases from tomato and Arabidopsis was aligned by ClustalW (UPGMA). The genes used were follows: SlCIPK1, 2, 8 and SlSOS2 (Table 1); AtCIPK1, At3g17510; AtCIPK2, At5g07070; AtCIPK8; At4g24400; AtSOS2 At5g35410; LeSNF1, AF143743; yeast SNF1, NP010765; SlSnRK2C, AB275884; AtSnRK2.8, AF41782; NtCDPK1, AF072908.
 (a)
(a) (b)
(b) (c)
(c)
Figure 2. The expression profiles of tomato CBL homologs, SlCIPK2, SlPP2C and SlUbi in tomato leaf and flower. (a) The expression levels of tomato CBLs; (b) The expression levels of SlCIPK2, SlPP2C and SlUbi. Semi-quantitative RT-PCR was carried out at indicated numbers of thermal; (c) A phylogenic tree of CBLs from tomato and Arabidopsis was aligned by ClustalW (UPGMA). The genes used were follows: SlCBL1, 2, 10 and SlSOS3 (Table 1); AtCBL1, At4g17615; AtCBL2, At5g55990; AtCBL10, At4g33000; AtCBL4/SOS3, At5g24270; tomato calmodulin (LeCaM), KTU3 Contig29992; Arabidopsis calmodulin (AtCaM1), At5g37780.
Total RNA was isolated from samples by the SDS/ phenol/LiCl method. For semi-quantitative RT-PCR, cDNA synthesis and PCR were performed with M-MLV reverse transcriptase (TaKaRa, Tokyo, Japan) and GoTaq Green Master Mix (Promega, Tokyo, Japan), respectively, according to the manufacturer’s manuals. Each 20 µL of PCR reaction mixture contained 10 mL of 2 × GoTaq mixture, 400 nM each 5’- and 3’-primer and 1 µL of tomato cDNA. The thermal cycle reaction was performed as follows: 94˚C for 2 min; 25 - 30 cycles (as indicated) of 94˚C for 10 s, 58˚C for 10 s, and 72˚C for 30 sec; 72˚C for 30 sec; and 4˚C for 99 min. Gene-specific primers for tomato CIPKs, tomato CBL homologs, a tomato putative Na+/H+ antiporter (SlSOS1) ortholog and tomato ubiquitin (SlUbi) are shown in Table 1. The PCR products separated in 1.5% agarose gels and then visualized by FluorChem (AlphaInnotech) after staining with ethidium bromide. The identity of these PCR products was confirmed by DNA sequencing.
2.3. Cloning and Construction of Plasmids
The full-length SlCIPK2 was amplified by PCR with cDNA prepared from flower-derived RNA, KOD plus polymerase (TOYOBO) and gene specific primers of SlCIPK2-FL-5 and SlCIPK2-3 (Table 1), according to the manufacturer’s manuals. After digesting by BamH I and Sal I, the DNA fragment was ligated between BamH I and Sal I of pMal-c, for expression of maltose-binding fusion protein (New England Biolabs, Massachusetts, USA). DNA fragments of SlCBL1, 2, 10, SlERF7, SlCBF1, SlABF4, SlAREB1 and SlPP2C were amplified by PCR with KOD plus DNA polymerase and sets of gene specific primers (Table 1). PCR products of SlCBLs1, 2, 10, SlERF7, SlCBF1, SlABF4 and SlAREB were digested by BamH I, and then inserted between BamH I and Sma I of pGEX4T-1 (Novagen, Darmstadt, Germany) in ligation. A PCR product of SlPP2C was cloned in pCR TOPO cloning kit (Invitrogen) and then subcloned into pGEX4T-1. Sequences of the cloned genes were confirmed by sequencing with an ABI Prism 310 DNA sequencer and Big Dye Terminator Cycle Sequencing Kit ver.1.1.
2.4. Expression and Purification of GSTand MBP-Fused Proteins
MBP-SlCIPK2 was purified with an amylose resin column according to the manufacturer’s (New England Biolabs) instruction manual. MBP-SlCIPK2 was expressed in E. coli BL21DE3 (Stratagene, La Jolla, CA, USA) containing pMal-c-SlCIPK2 by adding isopropyl b-D-1-thiogalactopyranoside (IPTG) at a final concentration of 0.1 mM when the OD600 of E. coli culture reached 0.5. After harvesting of E. coli cells (250 mL, 2 ´ YT medium) by centrifugation at 10,000 ´ g for 15 min, the E. coli pellet was resuspended in lysis buffer, containing 25 mM Tris·Cl [pH 7.5], 150 mM NaCl, 5 mM EDTA, 10% glycerol, 1% Triton X-100, 1 mM PMSF, and 0.1% b-mercaptoethanol, and then the cells were disrupted by sonication. After centrifugation of the mixture at 10,000 ´ g for 15 min at 4˚C, the resultant supernatant was applied to a column (1 cm ´ 2 cm) of amylose resins, and the column was washed with 20 mL of washing buffer containing 25 mM Tris-Cl [pH 7.4], 1 M NaCl, 0.5 mM EDTA, 0.1% Triton X-100, and 0.1% b-mercaptoethanol, then washed with elution buffer (2 mL), which is washing buffer supplemented with 10 mM maltose. The elutant was dialyzed overnight at 4˚C in 1 ´ TBS (25 mM Tris·HCl [pH 7.4], 150 mM NaCl) containing 0.5 mM EDTA, 0.1% Triton X-100, 0.1% b-mercaptoethanol, and 30% glycerol and then stored at –40˚C.
GST-fusion proteins were purified from E. coli extracts with glutathione CL4B beads, according to the manufacturer’s (GE Healthcare Life Sciences) instruction manual. Growth of E. coli cells containing pGEX plasmids, protein induction by IPTG and preparation of 10,000 ´ g supernatnat were performed as for MBP-fusion protein. The resultant supernatant was incubated with 200 mL of


Table 1. Primers used for RT-PCR analysis and construction of expression plasmids.
50% glutathione CL4B beads slurry (v/v) by rotating at 4˚C for 2 hours. After centrifugation at 3000 rpm for 1 min at 4˚C, the supernatants were discarded. The resultant GST-fusion protein bound beads were washed 5 times by centrifugation and adding washing buffer mention above. The purified GST-fusion proteins bound on glutathione bead were added with 30% glycerol at final concentration and then stored at –30˚C.
2.5. Immunoblot with Anti-CIPK Antibody, Anti-Tomato CBL4/SOS3 Antibody
After SDS-PAGE, separated polypeptides were electroblotted onto a PVDF membrane at 10 V cm–1 for 90 min in 25 mM Tris-base containing 0.05% SDS and 20% methanol) and then the PVDF membrane (Millipore) was blocked by incubating in 1 ´ TBS containing 3% skim milk (TBS-milk) for 60min. The PVDF membrane was incubated with anti-CIPK antibody (one to 2000 dilution [v/v]) or anti-tomato-SOS3 antibody (one to 2000 dilution [v/v]) [16] in TBS-milk containing 0.05% Tween 20 at room temperature for 2 hours. Subsequently, the PVDF membrane was incubated at room temperature for 60 min in TBS containing horseradish peroxidase (HRP)— conjugated protein G (GE Healthcare Bio-Sciences, Piscataway, NJ, USA) (one to 10,000 dilution [v/v]), and 0.05% Tween 20. The immunodecorated protein bands were visualized by an ECL Plus kit (GE Healthcare Bio-Sciences) and FluorChem Imaging Analyzer.
2.6. Preparation of Extracts from Plant Tissues
Frozen tissues (2 g) were homogenized on an ice-cooled motor in 6 mL of lysis buffer containing 20 mM 3-(Nmorpholino)-propanesulfonic acid (MOPS), 50 mM Na2-bglycerophosphate [pH 7.6], 10 mM EDTA, 5 mM EGTA, 30 mM NaF, 5 mM Na3VO4, 5% glycerol, 0.1% b-mercaptoethanol, 1 mM PMSF, 5 mM n-aminocaproic acid, 1 mM benzamidine, 1 mM Na-bisulfite, and 1 µg/mL leupeptin, with or without 1% Triton X-100. The resultant homogenates were centrifuged at 10,000 ´ g for 20 min at 4˚C. 100,000 ´ g supernatants (soluble fractions) and 100,000 ´ g pellets (microsomal fractions) were prepared by centrifugation at 100,000 ´ g for 60 min at 4˚C (P65A rotor and 55P-7 ultra-centrifugator, Hitachi Inc., Tokyo, Japan) with 10,000 ´ g supernatant fraction (without Triton X-100) described above. Protein concentrations in samples were measured by a Bio Rad protein assay kit (BioRad, Hercules, CA, USA) with bovine serum albumin (BSA) as a standard.
2.7. In Vitro Binding Assay
Ten micrograms each of GST and GST-fusion proteins bound on 20 mL of glutathione Sepharose beads (50% [v/v] slurry) in 1.5 mL of binding buffer containing 1 ´ TBS, 0.1% Triton X-100, and 0.1% b-mercaptoethanol, were added to 10 µg of purified MBP-SlCIPK2. After rotation at 4˚C for 2 hours, the GST-fusion protein-immobilized beads were briefly centrifuged at 5000 ´ g for 10 s and then washed 5 times with 1 mL of washing buffer containing 1 ´ TBS, 0.5 M NaCl, 0.1% Triton X-100, and 0.1% b-mercaptoethanol. After these washings, the precipitates with beads were separated by SDS-PAGE and the separated polypeptides were visualized by staining with Coomassie Brilliant Blue R250.
2.8. Construction of Green Fluorescence Protein Plasmids and Fluorescence Microscopy
A multicloning site containing Bgl II and Sal I was inserted at 3’-end of GFP(S65T) gene of an original GFP expression vector [17]. SlCIPK2 PCR product was digested with BamH I and Sal I as described in the construction of pGEX-SlCIPK2, and then was ligated into the Bgl II and Sal I site of pCaM35S-pro-GFP(S65T) (This study). The resultant pCaM35S-pro-GFP(S65T)-SlCIPK2 plasmid and the pCaM-pro-GFP(S65T) empty vector were introduced into onion epidermal cells with a particle bombardment (PDS-100/He, BioRad Ltd.) using 1.0- or 1.6-µm gold particles at 1100 psi. After incubation in Murasige-Skoog medium containing 3% sucrose for 24 hours, the onion cells were subjected to fluorescence microscopy analysis (ECLIPSE 80i, Nikon Ltd., Tokyo, Japan) with a filter system of BP480nm/DM550nm/LP510nm.
3. Results
3.1. A Flower Specific Expression of a Tomato CIPK Homolog
A phylogenic tree indicates that the tomato CIPKs and the corresponding Arabidopsis CIPKs formed a distant group from other SNF1-related kinases including SnRK I and SnRK II subfamilies and CDPK among the CDPKSNF1 superfamily, as expected (Figure 1(b)). A significant signal of SlCIPK2 was detected specifically in flower at 25 cycles, and the expression level of SlCIPK2 is the highest in flower than in other organs at 28 or even 31 cycles (Figure 1(a)). In contrast, SlCIPK1 were detected ubiquitously in all organs at 30 and 33 cycles (Figure 1(a)). At 27 cycles, the relative signals of SlCIPK8 and SlSOS2 were much weaker than those of SlCIPK1, while those were expressed in all organs. SlSOS1, expression of putative Na+/H+ antiporter, was also detected ubiquitously in all organs.
Next, a phylogenic tree of CBLs indicates relationships of tomato CBLs with the corresponding Arabidopsis CBLs by using tomato calmodulin (LeCaM) and AtCaM1 as outgroup (Figure 1(b)). Among CBL homologs, the expression of SlSOS3 were significant in leaf but not or marginal in flower at 27 and 30 cycles. The signals of SlCBL1 and SlCBL2 in flower were weaker than those in leaf at 27 cycles, but the difference of expression of SlCBL1 and SlCBL1 between leaf and flower was not apparent at 30 cycles. The signals of SlCBL10 showed no or marginal difference in transcript level between leaf and flower.
3.2. Localization of CIPK-Related Proteins and CBL-Related Proteins in Vaious Organs of Tomato
It was confirmed that the anti-cowpea CIPK antibody cross-reacts with MBP-SlCIPK2 but not with MBP only (Figure 3(a)). The anti-CIPK antibody can cross-react broadly with with CIPK-related proteins in higher plants, tomato, soybean, cowpea and Arabidopsis thaliana [16].
Immunoblot by the anti-CIPK antibody showed a significant signal in flower with a relative molecular mass of 55 kDa (Figure 3(b)) in the presence or the absence of Triton X-100. Weak signals were also detected in young seedling and leaf in the presence of Triton X-100 at about 55 kDa but no or marginal in the absence of Triton X-100. Immuno-reactive signals in root, stem and fruit were only very weak or faint. Next, the tissue specific profiles of the CIPK-related proteins in flower was determined by immunoblot using extracts, which were prepared by 10,000 ´ g centrifugation with homogenates from various tissues of tomato flower and plant in the presence of Triton X-100 (Figure 4(a)). The immunoreactive signals at 55-kDa were significant in stamen, and very low in petal, ovary and young fruit, but no or scarcely in leaf and petal.
To examine the intracellular localization of the flowerspecific CIPK-related polypeptide, ultracentrifugation was carried out with the 5000 ´ g supernatant fractions from homogenates of leaf and flower without Triton X-100. A significant signal was detected in 50,000 ´ g supernatant and 100,000 ´ g pellet in flower but not in 100,000 ´ g in flower. In contrast, there was no or marginal immunoreactive signal in samples of leaf (Figure 4(b), right). Several Arabidopsis CBLs are covalently bound to a myristoyl moiety on glycine at the N-terminus leading to recruitment of the CBL-CIPK complex on the microsomes [18]. Immunoblot revealed the SOS3-reated peptide at about 28 kDa in 100,000 ´ g pellets but not in 100,000 ´ g supernatants of leaf extracts (Figure 4(c), left). In contrast, there was no or marginal immunoreactive signals both in 100,000 ´ g super4C, right).
3.3. Interaction of SLCIPK2 with CBL Homologs and Transcriptional Factors
The deduced aimino acid sequence of SlCIPK2 indicates the presence of CBL-interacting domain called “NAF/ FISL motif” at 312 - 333 aa in the C-terminus [10]. Therefore, we used in vitro binding assay with a MBP-SlCIPK2 and GST-SlCBLs to examine whether SlCIPK2 associates with SlCBL1, 2 and 10, because it have been reported that the CBLs have different preference for potential counterparts of CIPKs [19]. In vitro binding assay of MBP-SlCIPK2 was performed using GST and GST-SlCBL1, 2 and 10. MBP-SlCIPK1 was detected in pull-downed samples with GST-SlCBL1, 2 and 10 but not with GST (Figure 5(a)). Interestingly, the presence or absence of Ca2+ in the binding buffer had marginal or no effect on the interaction of MBP-SlCIPK2 with GSTSlCBL1, 2 nor 10 (data not shown), although the predicted amino acid sequences of SlCBLs contain typical EF-hand motives that potentially bind with Ca2+. The Ca2+-dependent association of CIPKs with CBLs and the resultant activation is dependent on preferential combinations of
 (a)
(a) (b)
(b)
Figure 3. Detection of immunologically CIPK-related proteins in various organs of tomato. Recombinant SlCIK2 (a) and protein extracts prepared from seedling, root, stem leaf, flower and fruit (b) were analyzed by immunoblot with anti-CIPK specific antibody and ECL kit. Recombinant protein of MBP-SlCIPK2 and MBP were subjected to immunoblot by the antibody using indicated amounts of proteins. The 10,000 ´ g soluble fractions were prepared from crude extracts of leaves of each plants in the presence (right) or the absence (left) of 1% Triton X-100. Each 50 mg protein extracts per lane was subjected to SDS-PAGE (10% acrylamide gels) and electroblotted to PVDF membrane.
 (a)
(a) (b)
(b) (c)
(c)
Figure 4. Immunoblot of CIPK-related polypeptides and SOS3-related polypeptides in leaf and flower of tomato plant. Immunoblot using solubilized protein extracts prepared from leaf, sepal, petal, stamen, ovule and young fruit of tomato plant was carried out by anti-CIPK antibody (a). The soluble fraction and microsomal fraction prepared from leaf and flower were subjected to immunoblot by anti-CIPK antibody (b) and anti-SlSOS3 antibody (c). The 10,000 × g soluble fractions were prepared with lysis buffer containing 1% Triton X-100 from crude extracts of leaf and flower of 6-8-week-old tomato plant. Fractionation of 100,000 × g soluble fraction and 100,000 × g pellets was carried out at 100,000 × g for 60 min at 4˚C after preparation of 10,000 × g supernatants in the absence of Triton X-100. Each 50 μg protein per lane was subjected to SDS-PAGE (10% or 12.5% acrylamide gels) and immunoblot by anti-CIPK serum and anti-tomato CBL serum, respectively.
 (a)
(a) (b)
(b)
Figure 5. Interaction of SlCIPK2 with tomato CBL homologs and transcription factors. Pull-down assay with glutathionesepharose beads was carried out with MBP-SlCIPK2, GST, and GST-tomato CBL1, CBL2 and CBL10 in the presence of 1 mM CaCl2 (a). Recombinant proteins of tomato stress-responsive transcription factors and SlPP2C were subjected to in vitro binding assay of SlCIPK2 (b). Signals of MBP-SlCIPK2 co-precipitated with GST-fusion proteins are indicated by an arrow.
between individual CBLs and CIPKs [19]. For example, the Ca2+-independent interaction was observed between AtCBL4/SOS3 and AtCIPK24/SOS2 [15], as is the interaction between SlCIPK2 and SlCBL1, 2 and 10.
Recent studies revealed that AREB/ABFand AP2/ERFtype transcription factors are implicated in ABA and osmotic stress signaling [20,21]. Stress-activated SnRKs potentially phosphorylate those transcription factors [22- 26]. Therefore, in vitro interaction assay between SlCIPK2 and the tomato transcription factors, ERF7, CBF1, ABF4 and AREB1 was conducted. Furthermore, protein phosphatases type-2C (PP2C) appeaed to be involved in downregulation and dephosphorylation of CIPKs and SnRK2s and interacting with those specific CIPKs and SnRK2s [20,21]. Therefore, GST-SlPP2C was also subjected to the in vitro binding assay with SlCIPK2. MBP-SlCIPK1 was detected in pull-downed samples with GST-SlERF7, CBF1 and AREB1but not with GST nor GST-ABF4 (Figure 5(b)). There was no signal of MBP-SlCIPK2 in the pull-downed sample with GST-SlPP2C.
3.4. Subcellular Localization of GFP-Fused SLCIPK2
We investigated the distribution of GFP fluorescence of GFP-fused SlCIPK2 after transient expression into onion epidermal cells (Figure 6(a)). In these experiments, we observed a dispursed distribution of GFP-SlCIPK2 in the cytosol and a significant signal in whole nucleous. The pattern of subcellular distribution of GFP-SlCIPK2 distribution resembled to those of GFP alone, while the fluorescneces of GFP-SlCIPK were reltively weaker than those of GFP alone.
4. Discussion
The tomato and Arabidopsis CIPKs formed a monophyletic cluster separated from other clusters of SnRK1 groupe, LeSNF1 and yeast SNF1 [27], and of SnRK2 groupe, SlSnRK2C and AtSnRK2.8, while NtCDPK1 is located as outgroup [12,28]. SlCIPK2 has the highest similarity (62%) in amino acid sequence to that of AtCIPK2, and SlCIPK1, SlCIPK8 and SlSOS2 have the highest similarities (66, 70 and 72%) in amino acid sequences to those of AtCIPK1, AtCIPK8 and AtCIPK24/ SOS2, respectively. SlCIPK2 and AtCIPK2 are placed in the subclass including AtCIPK15/PKS3, which is involved in Ca2+-regulated ABA perception [29], while AtCIPK24/ SOS2, which is involved in Na+ excretion [30], and AtCIPK23, which is involved in regulation of K+ channels [14], belong to another subclass of CIPK family includeing a set of CIPK homologs involving ion regulation of ion transport. In contrast to the expression profiles of SlCIPK2 mRNA (Figure 1), other tomato CIPK homologs are broadly expressed in various tissues at relatively low levels, compared with that of SlCIPK2 in flower. The high expression level of SlCIPK2 in flower (Figure 1(b)) is consistent with the observation that significant immnoreactive signal of the CIPK-related polypeptide in flower and stamen (Figure 3(b)). It is well known that osmolyres, proline and sucrose, are highly accumulated in pollen and that severe drought and heat stress on pollen development have critical effects on the efficiency of the germination and fertilization. It is conceivable that SlCIPK2 functions on regulation of osmolyte-uptake mechanism in pollen. When expression patterns of SlCIPK1, 2 and 8 and SlSOS2 were examined using whole plant treated by various stresses, drought, NaCl, ABA, and chilling stress, no or marginal alteration in expression levels of those homologs were detected (data not shown).
SlSOS3 among tomato CBLs is preferentially expressed in leave, but less or scarcely in flower, while SlCBL10 was expressed in both leave and flower at the same levels. In vitro binding assay suggests that SlCBL1, 2 and 10 have potential to interact with SlCIPK2. In Arabidopsis, SOS3 appeared to interact with SOS2, which is involved in Na+-extrusion and salt tolerance of root. In tomato, SlSOS3 is induced under salt stress (Takahashi, Yuasa and Iwaya-Inoue, in preparation). Therefore, our observations and specific interaction between SOS3 and SOS2, previously reported, suggest that those CBL10 and in part, CBL1 and 2, possibly function at upstream of SlCIPK2 and that SlSOS3 may be involved in salt stress signaling but not in flower specific functions.
Results of RT-PCR and immunoblot indicate that the CIPK-related protein and the SOS3-related protein are expressed in flower and leaf alternatively, and that both proteins are preferentially localized in the microsomal fraction but not in the soluble fraction. The N-terminus of SlSOS3 contains a typical consensus motif, “H2N-MG-C-F-P-S-”, recognized as “H2N-M-G-X-X-X-(S/T)-” for myristoylation, suggesting that SlSOS3 is preferentially
 (a)
(a)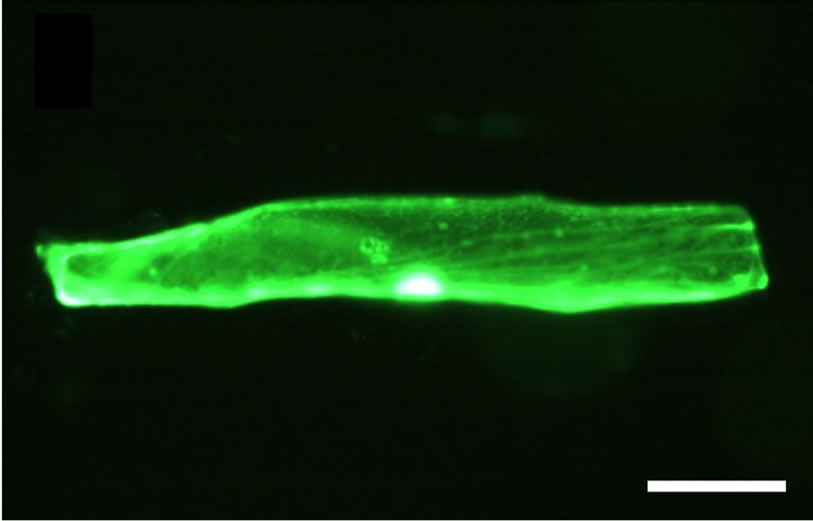 (b)
(b)
Figure 6. Green fluorescence proteins in onion epidermal cells introduced with plasmids expressing GFP-fused SlCIPK2 (a) and GFP alone (b). Onion epidermal cells were incubated on Murasige-Skoog medium containing 3% sucrose for 24 hours after gold particles coated with pCaM35S-pro-GFP(S65T)-fused SlCIPK2 and pCaM35S-pro-GFP(S65T) by particle gun. GFP expressed in onion epidermal cells was observed on a fluorescence microscopy. Bars indicated are 50 mm.
localized on the plasma membrane as SOS3 in Arabidopsis [31]. The SlCIPK2 has no apparent membrane associating domain in itself, while the flower specific CIPK was detected in the membrane fraction (Figure 4). Because that SlCBLl has the myristoyl motif “H2N-M-G-CF-N-S-” on its N-terminus, it is possible that SlCBL1 and/ or other lipid-modified CBL molecules recruit SlCIPK2 onto the membrane. In contrast to the result from subcellular fructionation (Figures 3(b) and 4(b)), the fluorescence of GFP-SlCIPK2 expressed in onion epidermis cells were detected in the cytosol and nucleous (Figure 6(a)). It was reported that distinct targeting signals located in the N-terminal domain of CBLs determine the spatially discrete localization of CBL/CIPK complexes via specific interaction set between CBLs and CIPKs [32]. Confocul microspcopic observation with coexpression of GFP-CIPKs and lipid-modified CBLs displayed significant fluorescence signals on the plasma membrane and/or the tonolplast, while expression of GFP-CIPKs alone showed the fluorescence signals in the cytosol and nucleous. As consistent to the previous stuides on CBL-CIPK, the subcellular fractionation and the fluorescence microscpic observation in the present study suggest that the membrane association of SlCIPK2 requires interaction with a specific CBLs possessing its membrane targeting signal.
There has been growing evidence that SnRK2s and SnRK3s participate coordinately in stress-stimulated gene expression by phosphorylating different sets of transcription factors in response to environmental stresses and hormonal signals, while previous studies have been focusing on regulation of ion transport on plasma membrane and in vitro binding assay showed that SlCIPK2 can associate with SlERF7, SlCBF1 and SlAREB. Recently, interaction of ABA signal-related PP2Cs, ABI1/ABI2 with SnRK2.6/AtSRK2E and SOS2 has been demonstrated [33,34]. While in vitro pull-down assay failed to detect association of SlCIPK2 with SlPP2C (Figure 5(b)), a possibility is not excluded that SlCIPK2 is regulated by other PP2C homologs.
The present study revealed unique features of SlCIPK2: 1) SlCIPK2 mRNA is expressed specifically in flower; 2) The CIPK-related protein is detected at a significant level in stamen; 3) The flower specific CIPK tightly associates with microsomal fraction of flower; 4) SlCIPK2 can interact to stress-responsive transcription factors. These observations suggest that SlCIPK is possibly involved in floral organ specific functions rather than salt stress signaling mediated by SlSOS3. It is necessary for clarifying the intracellular localization and the tissue specific expression of SlCIPK2 to perform immunofluorscence microscopic analysis and in-situ hybridization. While interaction of SlCIPK2 with CBL1, 2 and 10 was detected in in vitro binding assay, it is conceivable that the specific CBLs are required for association and function of SlCIPK2 on the membrane. It was reported that accumulation of proline and sucrose in pollen at high concentration is required for the development, maturation and desiccation. The osmolyte uptake mechanisms in pollen may be accounted for stress signaling cascades utilizing flowerspecific SlCIPK2 and CBLs which regulate solute transport and water permieability ofstamen and pollen cells. It remains to be clarified whether SlCIPK2 phosphorylates SlERF7, SlCBF1 and SlAREB, leading to activation of stress-inducible genes in flower, or whether it is involved in the direct regulation of transporters on vacuole and plasma membrane in floral tissues by phosphorylation along developmental process of pollen.
Increasing evidence suggests that CBL-CIPK complexes phosphorylate and activate various ion transporters, such as Na+/H+ exchangers and K+ channels [14,35], in vegetative organ such as root, stem and leave, whereas there were few reports about flower specific CIPK nor SnRK2. Therefore, identification of the activator or signaling regulated regulating SlCIPK2 in floral organ is important for improvement of stress tolerance of reproductive organ.
5. Acknowledgements
This work was supported in part by Grant-in-Aids from the Ministry of Education, Sports, Culture, Science and Technology of Japan, No. 19380022 (to T. Y.) and Salt Science Research Foundation, No. 0827 (to T.Y.).
REFERENCES
- N. Yabe, T. Takahashi and Y. Komeda, “Analysis of Tissue-Specific Expression of Arabidopsis thaliana HSP90- Family Gene HSP81,” Plant and Cell Physiology, Vol. 35, No. 8, 1994, pp. 1207-1219.
- C. Prasinos, K. Krampis, D. Samakovli and P. Hatzopoulos, “Tight Regulation of Expression of Two Arabidopsis cytosolic Hsp90 Genes during Embryo Development,” Journal of Experimental Botany, Vol. 56, No. 412, 2005, pp. 633-644. doi:10.1093/jxb/eri035
- R. Schwacke, S. Grallath, K. E. Breitkreuz, E. Stransky, H. Stransky, W. B. Frommer and D. Rentsch, “LeProT1, a Transporter for Proline, Glycine Betaine, and GammaAmino Butyric Acid in Tomato Pollen,” The Plant Cell, Vol. 11, No. 3, 1999, pp. 377-392. doi:10.1105/tpc.11.3.377
- K. Shinozaki and K. Yamaguchi-Shinozaki, “Molecular Responses to Dehydration and Low Temperature: Differences and Cross-Talk between Two Stress Signaling Pathways,” Current Opinion in Plant Biology, Vol. 3, No. 3, 2000, pp. 217-223. doi:10.1016/S1369-5266(00)80068-0
- J. L. Brewster, T. de Valoir, N. D. Dwyer, E. Winter and M. C. Gustin, “An Osmosensing Signal Transduction Pathway in Yeast,” Science, Vol. 259, No. 5102, 1993, pp. 1760-1763. doi:10.1126/science.7681220
- K. Ichimura, K. Shinozaki, G. Tena, J. Sheen, C. V. Giordano, A. Acevedo, A. L. Scopel and C. L. Ballare (MAPK Group), “Mitogen-Activated Protein Kinase Cascades in Plants: A New Nomenclature,” Trends in Plant Science, Vol. 7, No. 7, 2002, pp. 301-308. doi:10.1016/S1360-1385(02)02302-6
- J. Li, X. Q. Wang, M. B. Watson and S. M. Assmann, “Regulation of Abscisic Acid-Induced Stomatal Closure and Anion Channels by Guard Cell AAPK Kinase,” Science, Vol. 287, No. 5451, 2000, pp. 300-303. doi:10.1126/science.287.5451.300
- T. Umezawa, R. Yoshida, K. Maruyama, K. YamaguchiShinozaki and K. Shinozaki, “SRK2C, a SNF1-Related Protein Kinase 2, Improves Drought Tolerance by Controlling Stress-Responsive Gene Expression in Arabidopsis thaliana,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 101, No. 49, 2004, pp. 17306-17311. doi:10.1073/pnas.0407758101
- T. Yuasa, Y. Tomikubo, T. Yamauchi, A. Inoue and M. Iwaya-Inoue, “Environmental Stresses Activate a Tomato SNF1-Related Protein Kinase 2 Homolog, SlSnRK2C,” Plant Biotechnology, Vol. 24, No. 4, 2007, pp. 401-408. doi:10.5511/plantbiotechnology.24.401
- V. Albrecht, O. Ritz, S. Linder, K. Harter and J. Kudla, “The NAF Domain Defines a Novel Protein-Protein Interaction Module Conserved in Ca2+-Regulated Kinases,” The EMBO Journal, Vol. 20, No. 5, 2001, pp. 1051-1063. doi:10.1093/emboj/20.5.1051
- Y. Guo, U. Halfter, M. Ishitani and J.-K. Zhu, “Molecular Characterization of Functional Domains in the Protein Kinase SOS2 That Is Required for Plant Salt Tolerance,” The Plant Cell, Vol. 13, No. 6, 2001, pp. 1383-1400. doi:10.1105/tpc.13.6.1383
- E. M. Hrabak, C. W. Chan, M. Gribskov, J. F. Harper, J. H. Choi, N. Halford, J. Kudla, S. Luan, H. G. Nimmo, M. R. Sussman, M. Thomas, K. Walker-Simmons, J. K. Zhu and A. C. Harmon, “The Arabidopsis CDPK-SnRK Superfamily of Protein Kinases,” Plant Physiology, Vol. 132, No. 2, 2003, pp. 666-680. doi:10.1104/pp.102.011999
- V. Albrecht, S. Weinl, D. Blazevic, C. D’Angelo, O. Batistic, U. Kolukisaoglu, R. Bock, B. Schulz, K. Harter and J. Kudla, “The Calcium Sensor CBL1 Integrates Plant Responses to Abiotic Stresses,” Plant Journal, Vol. 36, No. 4, 2003, pp. 457-470. doi:10.1046/j.1365-313X.2003.01892.x
- J. Xu, H. D. Li, L. Q. Chen, Y. Wang, L. L. Liu, L. He and W. H. Wu, “A Protein Kinase, Interacting with Two Calcineurin B-Like Proteins, Regulates K+ Transporter AKT1 in Arabidopsis,” Cell, Vol. 125, No. 7, 2006, pp. 1347-1360. doi:10.1016/j.cell.2006.06.011
- U. Halfter, M. Ishitani and J.-K. Zhu, “The Arabidopsis SOS2 Protein Kinase Physically Interacts with and Is Activated by the Calcium-Binding Protein SOS3,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 97, No. 7, 2000, pp. 3735-3740. doi:10.1073/pnas.97.7.3735
- M. Imamura, T. Yuasa, T. Takahashi, N. Nakamura, M. P. S. H. Nang, S.-H. Zheng, K. Shimazaki and M. Iwaya-Inoue, “Isolation and Characterization of a cDNA Coding Cowpea (Vigna unguiculata (L.) Walp.) Calcineurin B-Like Protein Interacting Protein Kinase, VuCIPK1,” Plant Biotechnology, Vol. 25, No. 4, 2008, pp. 437-445.
- Y. Niwa, T. Hirano, K. Yoshimoto, M. Shimizu and H. Kobayashi, “Non-Invasive Quantitative Detection and Applications of Non-Toxic, S65T-Type Green Fluorescent Protein in Living Plants,” Plant Journal, Vol. 18, No. 4, 1999, pp. 455-463. doi:10.1046/j.1365-313X.1999.00464.x
- U. Kolukisaoglu, S. Weinl, D. Blazevic, O. Batistic and J. Kudla, “Calcium Sensors and Their Interacting Protein Kinases: Genomics of the Arabidopsis and Rice CBLCIPK Signaling Networks,” Plant Physiology, Vol.134, No. 1, 2004, pp. 43-58. doi:10.1104/pp.103.033068
- O. Batistic and J. Kudla, “Integration and Channeling of Calcium Signaling through the CBL Calcium Sensor/ CIPK Protein Kinase Network,” Planta, Vol. 219, No. 6, 2004, pp. 915-924. doi:10.1007/s00425-004-1333-3
- T. Hirayama and K. Shinozaki, “Perception and Transduction of Abscisic Acid Signals: Keys to the Function of the Versatile Plant Hormone ABA,” Trends in Plant Science, Vol. 12, No. 8, 2007, pp. 343-351. doi:10.1016/j.tplants.2007.06.013
- A. Wasilewska, F. Vlad, C. Sirichandra, Y. Redko, F. Jammes, C. Valon, N. F. dit Frey and J. Leung, “An Update on Abscisic Acid Signaling in Plants and More...” Molecular Plant, Vol. 1, No. 2, 2008, pp. 198-217. doi:10.1093/mp/ssm022
- Y. Uno, T. Furihata, H. Abe, R. Yoshida, K. Shinozaki, K. Yamaguchi-Shinozaki, “Arabidopsis Basic Leucine Zipper Transcription Factors Involved in an Abscisic AcidDependent Signal Transduction Pathway under Drought and High-Salinity Conditions,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 97, No. 21, 2000, pp. 11632-11637. doi:10.1073/pnas.190309197
- Y. Kobayashi, M. Murata, H. Minami, S. Yamamoto, Y. Kagaya, T. Hobo, A. Yamamoto and T. Hattori, “Abscisic Acid-Activated SNRK2 Protein Kinases Function in the Gene-Regulation Pathway of ABA Signal Transduction by Phosphorylating ABA Response Element-Binding Factors,” The Plant Journal, Vol. 44, No. 6, 2005, pp. 939- 949. doi:10.1111/j.1365-313X.2005.02583.x
- H. Choi, H. J. Park, J. H. Park, S. Kim, M. Y. Im, H. H. Seo, Y. W. Kim, I. Hwang and S. Y. Kim, “Arabidopsis Calcium-Dependent Protein Kinase AtCPK32 Interacts with ABF4, a Transcriptional Regulator of ABA-Responsive Gene Expression, and Modulates Its Activity,” Plant Physiology, Vol. 139, No. 4, 2005, pp. 1750-1761. doi:10.1104/pp.105.069757
- P. Agarwal, R. Arora, S. Ray, A. K. Singh, V. P. Singh, H. Takatsuji, S. Kapoor and A. K. Tyagi, “Genome-Wide Identification of C2H2 Zinc-Finger Gene Family in Rice and Their Phylogeny and Expression Analysis,” Plant Molecular Biology, Vol. 65, No. 4, 2007, pp. 467-485. doi:10.1007/s11103-007-9199-y
- C. P. Song, M. Agarwal, M. Ohta, Y. Guo, U. Halfter, P. Wang and J.-K. Zhu, “Role of an Arabidopsis AP2/ EREBP-Type Transcriptional Repressor in Abscisic Acid and Drought Stress Responses,” The Plant Cell, Vol. 17, No. 8, 2005, pp. 2384-2396. doi:10.1105/tpc.105.033043
- J. L. Celenza and M. Carlson, “Structure and Expression of the SNF1 Gene of Saccharomyces cerevisiae,” Molecular and Cellular Biology, Vol. 4, No. 1, 1984, pp. 54- 60.
- S. Ishida, T. Yuasa, M. Nakata and Y. Takahashi, “A Tobacco Calcium-Dependent Protein Kinase, CDPK1, Regulates the Transcription Factor Repression of Shoot Growth in Response to Gibberellins,” The Plant Cell, Vol. 20, No. 12, 2008, pp. 3273-3288.
- Y. Guo, L. Xiong, C.-P. Song, D. Gong, U. Halfter and J.-K. Zhu, “A Calcium Sensor and Its Interacting Protein Kinase Are Global Regulators of Abscisic Acid Signaling in Arabidopsis,” Developmental Cell, Vol. 3, No. 2, 2002, pp. 233-244. doi:10.1016/S1534-5807(02)00229-0
- Q. S. Qiu, Y. Guo, F. J. Quintero, J. M. Pardo, K. S. Schumaker and J.-K. Zhu, “Regulation of Vacuolar Na+/H+ Exchange in Arabidopsis thaliana by the Salt-OverlySensitive (SOS) Pathway,” Journal of Biological Chemistry, Vol. 279, No. 1, 2004, pp. 207-215. doi:10.1074/jbc.M307982200
- M. Ishitani, J. Liu, U. Halfter, C. S. Kim, W. Shi and J.-K. Zhu, “SOS3 Function in Plant Salt Tolerance Requires N-Myristoylation and Calcium Binding,” The Plant Cell, Vol. 12, No. 9, 2000, pp. 1667-1678. doi:10.1105/tpc.12.9.1667
- O. Batistic, R. Waadt, L. Steinhorst, K. Held and J. Kudla, “CBL-Mediated Targeting of CIPKs Facilitates the Decoding of Calcium Signals Emanating from Distinct Cellular Stores,” The Plant Journal, Vol. 61, No. 2, 2010, pp. 211-222.
- M. Ohta, Y. Guo, U. Halfter and J.-K. Zhu, “A Novel Domain in the Protein Kinase SOS2 Mediates Interaction with the Protein Phosphatase 2C ABI2,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 100, No. 20, 2003, pp. 11771-11776. doi:10.1073/pnas.2034853100
- R. Yoshida, T. Umezawa, T. Mizoguchi, S. Takahashi, F. Takahashi and K. Shinozaki, “The Regulatory Domain of SRK2E/OST1/SnRK2.6 Interacts with ABI1 and Integrates Abscisic Acid (ABA) and Osmotic Stress Signals,” Journal of Biological Chemistry, Vol. 281, No. 8, 2006, pp. 5310-5318. doi:10.1074/jbc.M509820200
- J.-K. Zhu, “Regulation of Ion Homeostasis under Salt Stress,” Current Opinion in Plant Biology, Vol. 6, No. 5, 2003, pp. 441-445. doi:10.1016/S1369-5266(03)00085-2
NOTES
*Corresponding author.