Journal of Modern Physics
Vol.08 No.08(2017), Article ID:77678,52 pages
10.4236/jmp.2017.88086
An Emergence of a Quantum World in a Self-Organized Vacuum―A Possible Scenario
Vladimir A. Manasson
Sierra Nevada Corporation, Irvine, California, USA

Copyright © 2017 by author and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: April 29, 2017; Accepted: July 14, 2017; Published: July 17, 2017
ABSTRACT
We have explored a model of vacuum self-organization based on dissipative dynamics and recurrent self-interactions. The initial state of the vacuum is assumed as self-interacting vacuum dust. The medium is dispersive and resembles dark-energy vacuum as described by general relativity. Beside self- diffusion, vacuum dust endowed with self-attraction, resembling Newton’s gravity. We explored what would happen with this medium when the strength of self-gravitation progressively increases. We observed a cascade of phase transitions. First transition occurs when self-attraction reaches the point when it can balance self-diffusion. A vortex-cellular structure emerges. Vortexes operate as self-sustained oscillators and tend to synchronize their dynamics. They form a synchronized network that possesses a universal time scale and, after zooming out, its structure acquires a form of fiber-bundle structure of electromagnetic field. With increasing self-gravitation strength, the system experiences another phase transition. The fiber-bundle structure becomes resembling that of weak nuclear field. Vacuum cells acquire spinorial dynamics. Electric charges emerge. When synchronized, the weakly interacting cells create lepton-like molecules. Oscillating charges in spinorial cells give a birth to current loops, which magnetic moment linked to the particle spin. During the next phase transition, the cell dynamics experiences another topological transformation, which is accompanied by creation of three color charges. The acquired fiber-bundle structure form resembles that of strong nuclear field. Synchronized strongly interacting vacuum cells create quark-like particles that carry color charges. We associate their complex synchronization patterns with particle flavors. We also explored statistical distributions of vacuum cells as functions of self-gravitation strength. We found that the distribution spectrum is essentially discrete, and the vacuum cells group around the states that we call super-attractive. Discrete cell distribution implies charge quantization. Synchronization transforms initial Boltzmann- like distribution into quantum-like distributions. During phase transitions, cell distributions experience transformations that can be encoded in the chemical potentials of the corresponding states. We found that chemical potentials apparently relate to the coupling constants and mixing angles and amplitudes in the standard model.
Keywords:
Elementary Particles, Standard Model, Quantized Charges, Quark Mixing, Self-Organization, Nonlinear Dynamics

1. Introduction
Before presenting the model, let me attract the reader’s attention to a few observations made by the author and other researchers [1] [2] [3] [4] .
1.1. Is Electron a Composite Particle?
Let us consider a thought experiment. An initially neutral vacuum volume experiences spontaneous polarization in a form of two charged clouds carrying charges 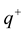 and
and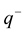 . Let us assume that the clouds interact with each other by pure electrostatic forces. In Figure 1, the clouds are represented by capacitor
. Let us assume that the clouds interact with each other by pure electrostatic forces. In Figure 1, the clouds are represented by capacitor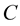 . After a while the clouds recombine and dissipate (radiate) the accumulated energy in the vacuum. The latter plays the role of a matched load with impedance
. After a while the clouds recombine and dissipate (radiate) the accumulated energy in the vacuum. The latter plays the role of a matched load with impedance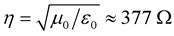 , where
, where 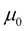 is the vacuum permeability and
is the vacuum permeability and 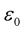 is the vacuum permittivity.
is the vacuum permittivity.

Figure 1. Cloud discharge circuit diagram. Charged clouds are represented by capacitor 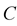 and the surrounding vacuum is represented by matched load
and the surrounding vacuum is represented by matched load .
.
Electrostatic energy stored in the clouds before discharge is
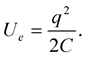 (1.1)
(1.1)
The corresponding relaxation time-constant is
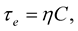 (1.2)
(1.2)
and the product 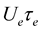 is C-independent and thus is independent of the cloud geometry
is C-independent and thus is independent of the cloud geometry
 (1.3)
(1.3)
If charged clouds represent a virtual electron-positron pair, 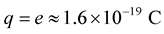 (electron charge) and product (1.3) is a physical constant with dimensions of action [energy × time]
(electron charge) and product (1.3) is a physical constant with dimensions of action [energy × time]
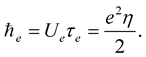 (1.4)
(1.4)
We use subscript 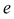 to distinguish
to distinguish 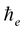 from the Planck constant
from the Planck constant 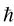 which has the same physical dimensions.
which has the same physical dimensions.
In quantum mechanics, the pair’s lifetime (dissipation time)  depends on their energy
depends on their energy  as
as
 (1.5)
(1.5)
If the total pair’s energy is due to the electrostatic forces, the two constants  and
and  would be of the same order (
would be of the same order ( ). However, they differ roughly by a factor of twenty! This is because the electrostatic forces contribute only a small part to the total particle energy.
). However, they differ roughly by a factor of twenty! This is because the electrostatic forces contribute only a small part to the total particle energy.
More important is the fact that the ratio  is a constant, and it is not only a physical constant but also a special number. With a good accuracy, it is
is a constant, and it is not only a physical constant but also a special number. With a good accuracy, it is
 (1.6)
(1.6)
where  is the Feigenbaum delta, a universal scaling factor describing complex transformations in dynamical systems belonging to a large universality class carrying the same name. The Feigenbaum universality was discovered decades ago [5] [6] [7] [8] . It has been observed both in theoretical models and experimentally in a number of different systems [9] - [14] , but, so far, not in particle physics.
is the Feigenbaum delta, a universal scaling factor describing complex transformations in dynamical systems belonging to a large universality class carrying the same name. The Feigenbaum universality was discovered decades ago [5] [6] [7] [8] . It has been observed both in theoretical models and experimentally in a number of different systems [9] - [14] , but, so far, not in particle physics.
In addition to electromagnetic, electrons/positrons interact via weak nuclear forces. Their relative “strength” is provided by ratio of two physical constants  (
( [15] [16] is the weak coupling constant and
[15] [16] is the weak coupling constant and  [15] [16] is the electromagnetic coupling constant better known as the fine-structure constant). Curiously (and we believe not by accident) the ratio can also be expressed via the Feigenbaum delta
[15] [16] is the electromagnetic coupling constant better known as the fine-structure constant). Curiously (and we believe not by accident) the ratio can also be expressed via the Feigenbaum delta
 (1.7)
(1.7)
Weak nuclear forces operate at very short distances only. It is unlikely that the two separated charged particles interact via these forces. It is more plausible to assume that the weak forces act inside the particles, i.e. that electron is a composite particle, and the weak forces keep its parts together.
1.2. Coupling Constants as Ordered Set of Numbers
Our encounters with the Feigenbaum constant are not completed. Another physical constant, the electroweak mixing angle  known as the Weinberg angle can also be expressed via
known as the Weinberg angle can also be expressed via . The angle is linked to the inverse
. The angle is linked to the inverse  ratio as
ratio as
 (1.8)
(1.8)
The fine structure constant has been the subject of numerous speculations and its origin remains an unsolved puzzle. It is defined as [16]
 (1.9)
(1.9)
Using this definition and (1.6) one can find that [1] [3] [4]
 (1.10)
(1.10)
From (1.7) and (1.9), the weak coupling constant is
 (1.11)
(1.11)
If one extends this progression toward the strong coupling constant , (s)he finds that
, (s)he finds that
 (1.12)
(1.12)
that is in the range of low-energy values recommend by the Particle Data Group [16] .
Thus we observe that the coupling constants of strong, weak, and electromagnetic interactions can be expressed as three consecutive powers of the Feigenbaum delta (up to a multiplier ):
):
 (1.13)
(1.13)
where  respectively.
respectively.
1.3. Quark Mixing Amplitudes and Hidden Symmetry of CKM-Matrix
Yet another example of quantum numbers that can be approximated by powers of  is a set of the quark mixing amplitudes
is a set of the quark mixing amplitudes , where
, where  and
and  denote different quark flavors. The
denote different quark flavors. The  value set is shown below as a
value set is shown below as a  Cabibbo- Kobayashi-Maskawa (CKM) matrix [17]
Cabibbo- Kobayashi-Maskawa (CKM) matrix [17]
 (1.14)
(1.14)
CKM-matrix elements can also be approximated as ,
, . The approximate
. The approximate  -matrix is
-matrix is
 (1.15)
(1.15)
The expressions under the square roots have form that preserves raw/column unitarity. In future discussion, due to the smallness of ,
,  , and
, and , we neglect their inputs in diagonal elements, and set the latter equal to 1 as shown in (1.16):
, we neglect their inputs in diagonal elements, and set the latter equal to 1 as shown in (1.16):
 (1.16)
(1.16)
 -matrix elements constitute an interesting pattern. First, the matrix possesses a diagonal symmetry. Second, each row/column comprises two out of three powers
-matrix elements constitute an interesting pattern. First, the matrix possesses a diagonal symmetry. Second, each row/column comprises two out of three powers ,
,  , and each time the missed power is a different one. Third,
, and each time the missed power is a different one. Third,  -matrix elements represent a subset of a
-matrix elements represent a subset of a  matrix where all powers are present:
matrix where all powers are present:
 (1.17)
(1.17)
In (1.17), missing elements fill the third row and third column. The term in parentheses in front of the matrix restores the raw/column unitarity.
We see that coupling constants and quark mixing amplitudes can be approximated as ordered sets of powers of the Feigenbaum delta. Could this be just a curious coincidence? We believe it is not.
1.4. Why Self-Organization?
The standard model does not explain why the coupling constants and mixing angles are constants and why they possess the values which we observe in experiment. Therefore, the possibility of expressing all of them through a single number, even approximately, is appealing, and more because this number is a universal constant. Why has  not found a place in the standard model of particle interactions?
not found a place in the standard model of particle interactions?
A possible explanation is that the two belong to the different frameworks. The dynamical systems with a shared feature of transition to chaotic dynamics through cascades of period-doubling bifurcations with the Feigenbaum delta  as the scaling factor belong to dissipative framework. In contrast, the standard model belongs to the Hamiltonian systems essentially based on the conservation principles.
as the scaling factor belong to dissipative framework. In contrast, the standard model belongs to the Hamiltonian systems essentially based on the conservation principles.
Although, the theory predicts that cascades of period-doubling bifurcations can also occur in some nonintegrable conservative systems [12] [14] [18] [19] [20] [21] [22] , this has not been confirmed in experiment. Moreover, in this case the scaling factor  essentially differs from
essentially differs from , observed in dissipative systems.
, observed in dissipative systems.
Conservation in Hamiltonian systems is at odds with emergence. Strict time translation symmetry implies that there should neither be a big bang nor “small bangs/crunches” linked to the births/deaths of elementary particles. Conservation rather preserves status quo.
In contrast, if a dissipative system is far from thermodynamic equilibrium, it may exhibit an entire gamut of emergent phenomena [23] [24] [25] [26] [27] . Dissipative frameworks is more appropriate for studying emergent phenomena.
In this paper we propose a new model that describes emergence of quantum phenomena and elementary particles. We adopt a conjecture that  is relevant to particle physics. We assume that the “primordial soup” where particles emerge and decay is a dissipative self-organized medium which we call vacuum.
is relevant to particle physics. We assume that the “primordial soup” where particles emerge and decay is a dissipative self-organized medium which we call vacuum.
One of the requirements for a dissipative system to be self-organized is that the system should be far from thermodynamic equilibrium. Numerous experiments witness that the vacuum is such a medium. It is permeated by numerous energy flows, at all levels, from quantum fluctuations to running galaxies.
Although this is not an exact parallel, Bénard flows (a network of vortices in nonequilibrium fluid) [28] may serve as a visual addition to the proposed model. This is supported by experiments where Bénard cells exhibit period-doubling bifurcations and the Feigenbaum universality [29] [30] [31] [32] [33] . Bénard cells are vortices that in essence can be described as self-sustained oscillators and tend to synchronize their dynamics, a phenomenon playing a principal role in the proposed model.
As any imitation, our model captures the most essential features only. We believe that by pursuing this approach, we may enrich our knowledge of the elementary particles and quantum fields.
2. Cellular Dynamics
2.1. From Vacuum Dust to Vacuum Cells
A simple example of spatial self-organization producing a quantized pattern is a randomly distributed iron filings placed between electromagnet poles. After field is turned on, the particles form a discrete field-line pattern. It happens under action of recurrent positive-feedback loops: small deviations from the uniform filings distribution create spots of excessive magnetic field; these spots attract more particles; the additional particles increase the local magnetic field; and so on. The filings redistribution ends when friction balances positive-feedback forces. The stronger is the external magnetic field the narrower are the field lines. The original symmetry breaks spontaneously. The locations of the field lines are arbitrary and depend on the initial filings fluctuations. Formation of the discrete field-line pattern is a phase transition. It can be seen as a “competition” among spatial positions for accumulating maximal number of particles, and the “fortunate” spots that initially have more particles are the winners. This competition resembles the Darwinian competition among the species.
More complex media can produce spatial-temporal patterns [34] [35] [36] [37] [38] . Bénard cells [28] emerging in heated liquids are among those.
In this paper, we assume that elementary particles emerge in vacuum as products of its self-organization. We assume that vacuum consists of ever moving vacuum dust. The medium is active. The dust particles attract each other. Self-attraction is nonlinear, i.e. the denser is the dust, the stronger is self-attraction. Self-attraction competes with self-diffusion. They represent respectively a positive and negative feedbacks. Stable dynamical patterns emerge when self-attraction is balanced by self-diffusion.
The vacuum dust is in a permanent motion. The state space comprises infinitively many different modes. Luckily, the system is dissipative, which means that the modes fade out and, generally speaking, with different rates. Therefore, for many applications, it is sufficient to consider only a few principal modes with slowest dissipation rates [9] [11] [12] .
We assume that most relevant modes represent vortices, local circular flows. If vortex life is long enough to perform at least several rotations, we call it a vacuum cell, or just cell. Dust migration connects vortices and forces them to synchronize in a manner of synchronization between self-sustained oscillators [39] [40] [41] [42] [43] .
We also focus on intracellular radial flows. We associate them with generic charges. We assign positive sign to convergent flows, negative sign to divergent flows, and zero to pure circular flows (Figure 2). Later in the paper, depending on cell topology, we categorize generic charges as electric charges and color charges.

Figure 2. Charge polarity assignment (directions of radial flows are opposite to the directions of field lines adopted in electrodynamics).
We define  as a probability that, in a given cell at a given time, the effective radial flow is converging. Using this convention, a cell is positively charged if
as a probability that, in a given cell at a given time, the effective radial flow is converging. Using this convention, a cell is positively charged if , it is negatively charged if
, it is negatively charged if , and it is neutral if
, and it is neutral if . Positive feedback (self-attraction) increases probability
. Positive feedback (self-attraction) increases probability  with time. Negative feedback (self-diffusion) decreases probability
with time. Negative feedback (self-diffusion) decreases probability  with time.
with time.
We assume that cell evolution under combined positive- and negative-feed- back can be effectively represented by a one-dimensional discrete iterated map
 (2.1)
(2.1)
where term  represents positive feedback, term
represents positive feedback, term  represents negative feedback, and parameter
represents negative feedback, and parameter  is a measure of the medium amplification.
is a measure of the medium amplification.
Discrete points  represent continuously evolving trajectories sampled stroboscopically at cross-sections with Poincaré plane (Figure 3). Fixed points
represent continuously evolving trajectories sampled stroboscopically at cross-sections with Poincaré plane (Figure 3). Fixed points  represent single-loop (Figure 3(b)) or multi-loop (Figure 3(c)) attractors and respectively are single-valued or multiple-valued.
represent single-loop (Figure 3(b)) or multi-loop (Figure 3(c)) attractors and respectively are single-valued or multiple-valued.
Dissipative evolution trajectories in the state space represent spirals (Figure 3(a)). The “stroboscopic” samplings are taken at time instants , when the trajectory is crossing the selected Poincaré plane.
, when the trajectory is crossing the selected Poincaré plane.

Figure 3. (a) Stroboscopic sampling of evolution trajectories; (b) single-loop attractor provides single-valued fixed point; (c) double-loop attractor provides double-valued fixed point.
Map (2.1) is known as logistic map [13] [14] . It obviously does not cover the complex turbulent picture of the medium in all nuances, but we assume that it is sufficient for the task. We justify this simplification by the following reasoning:
a) Map (2.1) describes dissipative dynamics
b) It accounts for the competition between positive and negative feedback in a simple form
c) It represents a wide class of theoretically explored and experimentally observed dynamical systems with the Feigembaum universality
d) It encompasses period-doubling bifurcations with the scaling factor .
.
e) Unlike one-dimensional continuous-time differential equations, it is flexible enough to describe topologically complex trajectories (like those shown in Figure 3(c)).
We define generic cellular charge  as
as
 (2.2)
(2.2)
Its domain  is inherited from probability
is inherited from probability  and is not typical for charges (
and is not typical for charges ( ). We could rescale
). We could rescale  by using an appropriate map or equation, such as this one
by using an appropriate map or equation, such as this one
 (2.3)
(2.3)
However, since in this paper we are looking only for the roots of charge quantization, its domain size is irrelevant. Therefore, we remain with  without further makeovers.
without further makeovers.
For mostly aesthetic reasons, we replace amplification parameter  with control parameter
with control parameter 
 (2.4)
(2.4)
with the corresponding changes in the domain size .
.
The lower end,  , is a critical point. Below this point,
, is a critical point. Below this point,  (
( ), the medium loses amplification, feedback is pure negative, balance of forces is broken, and self-organization fails.
), the medium loses amplification, feedback is pure negative, balance of forces is broken, and self-organization fails.
Initially, we study cell dynamics at different but fixed B-values. Later, we introduce a mechanism of vacuum self-regulation when the medium controls B-values by itself.
With the above introduced variables, the charge evolution is described by the iterated map
 (2.5)
(2.5)
where
 (2.6)
(2.6)
is iteration function.
In any dynamical system time plays most important role. Time is meaningless without clocks. However, self-organized vacuum cells are in essence self-sus- tained oscillators and can be used to clock the local time by themselves. As time unit, it is natural to use iteration intervals  between any two consecutive crossings of a charge trajectory with one of the Poincaré planes selected for this goal. If different cells operate asynchronously, each of them clocks their local time with its own pace. When the cells synchronize, interval
between any two consecutive crossings of a charge trajectory with one of the Poincaré planes selected for this goal. If different cells operate asynchronously, each of them clocks their local time with its own pace. When the cells synchronize, interval  acquires a global character and may serve as a time-standard for the entire synchronized network.
acquires a global character and may serve as a time-standard for the entire synchronized network.
2.2. Bifurcation Diagram. Zones of Stability. Special Points
Quadratic maps (including logistic map) are well explored (see, for example [5] - [14] ). For most researchers, they are particularly interesting as a paradigm of systems evolving toward chaotic dynamics. Ironically, we are more interested in the patterns of asymptotic stability, when a system asymptotically converges to some dynamically stable state.
Asymptotic stability varies with B. Continuous B-intervals where cell possess asymptotic stability we call zones of stability or just zones. On bifurcation diagram (Figure 4) which depicts fixed charge values  vs.
vs. , they are desig- nated as
, they are desig- nated as , where
, where  is the zone number. The diagram name came from bifurcations, the points where cell dynamics experience deep changes including changes in the attractor topology. We designate left and right bifurcations at the ends of each zone of stability respectfully as
is the zone number. The diagram name came from bifurcations, the points where cell dynamics experience deep changes including changes in the attractor topology. We designate left and right bifurcations at the ends of each zone of stability respectfully as  and
and . Bifurcations are points of unstable equilibrium. At bifurcations, number of attractor loops (number of branches in the diagram) doubles. Within each zone, the system is asymptotically stable. When
. Bifurcations are points of unstable equilibrium. At bifurcations, number of attractor loops (number of branches in the diagram) doubles. Within each zone, the system is asymptotically stable. When  reaches the Feigenbaum point,
reaches the Feigenbaum point,  , the system becomes chaotic.
, the system becomes chaotic.
In this paper we explore only the first three zones,  through
through , shown in Figure 4. We associate them respectfully with electromagnetic, weak, and strong nuclear interactions.
, shown in Figure 4. We associate them respectfully with electromagnetic, weak, and strong nuclear interactions.

Figure 4. Bifurcation diagram depicts fixed charges  as function on parameter
as function on parameter . Zones
. Zones  are marked by horizontal brackets, bifurcations
are marked by horizontal brackets, bifurcations  and
and  by black vertical dashed lines, superattractors
by black vertical dashed lines, superattractors  by red vertical dashed lines. Small color circles depict superattractor charges.
by red vertical dashed lines. Small color circles depict superattractor charges.
Later, we also explore interval , which we call “the dark zone” and designate as
, which we call “the dark zone” and designate as , where vacuum self-organization fails.
, where vacuum self-organization fails.
In , there is only one branch on the bifurcation diagram, which means that the cell attractors represent single loops, and fixed charges
, there is only one branch on the bifurcation diagram, which means that the cell attractors represent single loops, and fixed charges  are single- valued. In the higher zones, the number of branches doubles with each bifurcation, i.e. the attractors possess several loops, and fixed charges are multi-valued;
are single- valued. In the higher zones, the number of branches doubles with each bifurcation, i.e. the attractors possess several loops, and fixed charges are multi-valued; , where
, where  is the winding number (or number of the attractor loops). For example, attractor in Figure 3(b) has
is the winding number (or number of the attractor loops). For example, attractor in Figure 3(b) has , and attractor in Figure 3(c) has
, and attractor in Figure 3(c) has .
.
In zone , the winding number is
, the winding number is
 (2.7)
(2.7)
Full cell rotation period  comprises all different attractor loops and is
comprises all different attractor loops and is
 (2.8)
(2.8)
where  is the iteration time interval in zone
is the iteration time interval in zone .
.
For multi-loop attractor cells with  we reserve a special name. We call them spinorial cells [44] . They play a special role in toy-leptons and toy-quarks (we call them q-arcs) building blocks.
we reserve a special name. We call them spinorial cells [44] . They play a special role in toy-leptons and toy-quarks (we call them q-arcs) building blocks.
Superattractors are points of the highest asymptotic stability. We designate them as . In
. In , the superattractor is located exactly at the zone center. In all other zones, superattractors are slightly shifted from the centers. In
, the superattractor is located exactly at the zone center. In all other zones, superattractors are slightly shifted from the centers. In , the superattractor is located at the zone edge. We call intervals between superattractors
, the superattractor is located at the zone edge. We call intervals between superattractors  and left
and left  or right
or right  bifurcations semi-zones and designate them as
bifurcations semi-zones and designate them as  and
and .
.
Small circles at superattractors in Figure 4 depict superattractor charges. Zero charges are shown by white circles, and nonzero charges are shown by color circles (purple in , and red, green, and blue in
, and red, green, and blue in ).
).
B-values for special points on the bifurcation diagram are listed in Table 1.

Table 1. B-values for special points in zones  through
through .
.
With each bifurcation, zone size  decreases approximately as geometric progression .
decreases approximately as geometric progression .
 (2.9)
(2.9)
where scaling factor  quickly converges to the Feigenbaum delta
quickly converges to the Feigenbaum delta 
2.3. Cellular Evolution in Z1. Uncertainty Intervals
 cells are the simplest in the model and possess single-loop attractors. In this section, we explore how the charge asymptotic stability depends on a fixed parameter
cells are the simplest in the model and possess single-loop attractors. In this section, we explore how the charge asymptotic stability depends on a fixed parameter  in
in . We will study cell evolution toward the attractors for different
. We will study cell evolution toward the attractors for different  values.
values.
We assume that cells evolution may start from any initial charge value  between 1 and −1. A few examples of trajectories
between 1 and −1. A few examples of trajectories  are shown in Figure 5. All trajectories converge toward their fixed points. Trajectories with different parameters
are shown in Figure 5. All trajectories converge toward their fixed points. Trajectories with different parameters  are represented by different colors. They converge to different fixed points. However, all trajectories belonging to the same B-value converge to the same fixed point
are represented by different colors. They converge to different fixed points. However, all trajectories belonging to the same B-value converge to the same fixed point .
.
At the beginning of the cell evolution, the trajectories converging to one attractor entangle with trajectories converging to a different attractor. The attractors can be resolved only after some elapsing time. The longer is the elapsing time, the better is the attractor resolution.

Figure 5. Charge evolution trajectories  for three parameters
for three parameters  (shown in three colors).
(shown in three colors).
The entanglement and asymptotic behavior of charge trajectories have a qualitative analog in quantum mechanics in a form of the time-energy uncertainty relation―the longer a system is located at some state the better the state energy (fixed point) can be discriminated. This comparison implies that quantum-me- chanical eigenstates possess asymptotic stability thus quantum-mechanical systems are in essence dissipative, which is in agreement with the dissipative character of the wave-function collapses.
To quantitatively compare asymptotic stabilities of different attractors, we need to have a measure of the asymptotic stability. First, we define a charge uncertainty interval  for a given trajectory as
for a given trajectory as
 (2.10)
(2.10)
where  is the iteration step number after beginning of the cell evolution. Then we define parameter
is the iteration step number after beginning of the cell evolution. Then we define parameter  as a dimensionless slope of
as a dimensionless slope of  trajectory depicted in logarithmic scale
trajectory depicted in logarithmic scale
 (2.11)
(2.11)
After a few iteration steps (see Figure 6), the slopes exponentially converge to their fixed values
 (2.12)
(2.12)
From Figure 6, one can see that the slopes depend only on parameter .
.

Figure 6. Evolution of uncertainty intervals  (in logarithmic scale) for trajectories shown in Figure 5.
(in logarithmic scale) for trajectories shown in Figure 5.
Slope  plays the role of the Lyapunov exponent. It is a measure of the cell asymptotic stability. Within the zone, it is always negative
plays the role of the Lyapunov exponent. It is a measure of the cell asymptotic stability. Within the zone, it is always negative , approaches its minimum
, approaches its minimum  at the super attractor, and reaches its maximum
at the super attractor, and reaches its maximum  at the zone edges (bifurcations).
at the zone edges (bifurcations).
2.4. Spinorial Cells
We define spinorial cells as those with attractors having more than one loop ( ).
).
They serve as particle building blocks. We encounter two types of spinorial cells, d-cells ( from double-loop attractor) with winding number
from double-loop attractor) with winding number  (Figure 3(c)), and q-cells (
(Figure 3(c)), and q-cells ( from quadruple-loop attractor) with
from quadruple-loop attractor) with .
.
D-cells live in zone . An example of Z2-cell charge evolution trajectory is shown in Figure 7 by green seesaw line. In the limit
. An example of Z2-cell charge evolution trajectory is shown in Figure 7 by green seesaw line. In the limit , d-cell oscillates between two fixed values of the double-valued fixed points. We can split the single-step trajectory on two double-step trajectories, each with its own single- valued fixed point. The odd-step trajectory
, d-cell oscillates between two fixed values of the double-valued fixed points. We can split the single-step trajectory on two double-step trajectories, each with its own single- valued fixed point. The odd-step trajectory  is shown by the black dotted line. It converges toward fixed value
is shown by the black dotted line. It converges toward fixed value . The even-step trajectory
. The even-step trajectory  is shown by the red dotted line. It converges toward
is shown by the red dotted line. It converges toward .
.
For double-step trajectories we can explore the cell asymptotic stability in the same way as we did it for  cells.
cells.

Figure 7. Example of 𝑑-cell charge evolution.
For each double-step trajectory, we define a charge uncertainty interval as
 (2.13)
(2.13)
Remarkably, both uncertainty intervals  and
and  exponentially decay with the same rates (Figure 8), i.e. with the same slopes
exponentially decay with the same rates (Figure 8), i.e. with the same slopes

Figure 8. Example of evolution of 𝑑-cell uncertainty interval.
 (2.14)
(2.14)
Double-step evolution trajectories are described by twice-iterated functions  and
and  sampled at alternating time instants
sampled at alternating time instants
 (2.15)
(2.15)
Q-cells live in zone . Their attractors have four loops, and their fixed points are four-valued. An example of Z3-cell charge evolution trajectory is shown in Figure 9 by green seesaw line. It rotates around four dotted lines (shown in black, red, green, and blue) each depicting a four-step trajectory,
. Their attractors have four loops, and their fixed points are four-valued. An example of Z3-cell charge evolution trajectory is shown in Figure 9 by green seesaw line. It rotates around four dotted lines (shown in black, red, green, and blue) each depicting a four-step trajectory,  , where
, where  is the step number within a single period. The color states are ordered. The colors always come in the same sequence: ∙∙∙ → black → red → green → blue → black→ ∙∙∙. Due to the time irreversibility this order cannot be inverted. This order and irreversibility are crucial in discussion of quark (or more correctly, q-arc) mixing angles.
is the step number within a single period. The color states are ordered. The colors always come in the same sequence: ∙∙∙ → black → red → green → blue → black→ ∙∙∙. Due to the time irreversibility this order cannot be inverted. This order and irreversibility are crucial in discussion of quark (or more correctly, q-arc) mixing angles.

Figure 9. Example of q-cell charge evolution.
Four-step charge uncertainty intervals  are defined as
are defined as
 (2.16)
(2.16)
Their typical evolution is shown in Figure 10. As in the previous case, all of them have the same slopes .
.
Four-step iteration functions  are four-times iterated the original function
are four-times iterated the original function  sampled at different time instants
sampled at different time instants
 (2.17)
(2.17)
As in the previous case, initial charge  of the first trajectory can be chosen arbitrarily, while the initial charges of the other trajectories
of the first trajectory can be chosen arbitrarily, while the initial charges of the other trajectories  are images of
are images of  after the corresponding number of iterations.
after the corresponding number of iterations.

Figure 10. Example of evolution of q-cell uncertainty interval.
The multi-step trajectories have single-valued fixed points and are more suitable for analysis. Examples of ,
,  and
and , for a randomly selected B-values from each of
, for a randomly selected B-values from each of ,
,  , and
, and  zones, are shown in Figure 11.
zones, are shown in Figure 11.

Figure 11. Examples of ,
,  and
and  for randomly selected
for randomly selected  values.
values.
In each semi-zone, asymptotic stability depends on how far the function local extrema located from the super attractor in B-space. We define the distance between the cell attractor ( ) and the superattractor
) and the superattractor  as
as
 (2.18)
(2.18)
Because different semi-zones have different sizes, to compare asymptotic stability in different zones, we use reduced distance , which we define as
, which we define as
 (2.19)
(2.19)
where the denominator is the size of the corresponding semi-zone,  is the zone number,
is the zone number,  is superattractor,
is superattractor,  is the left bifurcation, and
is the left bifurcation, and  is the right bifurcation. The reduced distance domain is the same in all zones
is the right bifurcation. The reduced distance domain is the same in all zones  . For all superattractors
. For all superattractors , and for bifurcations
, and for bifurcations .
.
Before comparing asymptotic stabilities in different zones, we also need to bring them to the same time scale. We assume, that after cells become synchronized, they all have the same period 
 (2.20)
(2.20)
Correspondingly, their iteration intervals in zones  are
are
 (2.21)
(2.21)
With these assumptions, we extend definition of parameter  given by (2.11) for all zones
given by (2.11) for all zones  as
as
 (2.22)
(2.22)
After a few iteration steps (see Figure 10) the trajectories almost reach their fixed points and the slopes can be determined as
 (2.23)
(2.23)
Simulated dependences  in six semi-zones, from
in six semi-zones, from  through
through , are shown in Figure 12 by different symbols. All six are similar to each other and can be approximated as a logarithmic function (red solid curve)
, are shown in Figure 12 by different symbols. All six are similar to each other and can be approximated as a logarithmic function (red solid curve)
 (2.24)
(2.24)

Figure 12. Calculated functions  for six semi-zones (
for six semi-zones ( through
through ) are represented by different symbols. Solid red line depicts function
) are represented by different symbols. Solid red line depicts function .
.
2.5. Energy and Chemical Potentials
We define cell energy as
 (2.25)
(2.25)
 has physical dimensions of frequency. It inherits many features of parameter
has physical dimensions of frequency. It inherits many features of parameter . For example, we can use it as a measure of cell asymptotic stability. Excluding bifurcations, where
. For example, we can use it as a measure of cell asymptotic stability. Excluding bifurcations, where , it is always negative and takes the most negative values at the superattractors. It is customary to associate negative energies with the bound states. In our case, a vacuum cell is bound by the dust self- attraction forces.
, it is always negative and takes the most negative values at the superattractors. It is customary to associate negative energies with the bound states. In our case, a vacuum cell is bound by the dust self- attraction forces.
Using (2.24) and (2.25), we obtain that energy depends on parameter  as
as
 (2.26)
(2.26)
If we neglect the small differences between the left and right semi-zones, from (2.9) we obtain that
 (2.27)
(2.27)
From (2.26) and (2.27), we obtain that energy depends on  in all zones as
in all zones as
 (2.28)
(2.28)
where , and
, and  is a parameter that we associate with chemical potential (internal cellular energy) in the corresponding zone and is defined as
is a parameter that we associate with chemical potential (internal cellular energy) in the corresponding zone and is defined as
 (2.29)
(2.29)
Cellular energy  as function on parameter
as function on parameter  is shown in Figure 13. It has a form of three connected potential wells, each covering its own zone, and having deep minima at the superattractors.
is shown in Figure 13. It has a form of three connected potential wells, each covering its own zone, and having deep minima at the superattractors.

Figure 13.  in
in  through
through  (bottom) and bifurcation diagram (top).
(bottom) and bifurcation diagram (top).
The well shapes are almost identical. Z2-well and Z3-well can be obtained from Z1-well by horizontal and vertical translations (see (2.30)) as it is shown in Figure 14. Vertical shifts are equal to the chemical potential differences in the corresponding zones.
 (2.30)
(2.30)

Figure 14. Directly calculated Z2-well (blue) and Z3-well (red), and translated copies of Z1-well (black) according to (2.30).
2.6. Partition Function. Vacuum Self-Regulation. Discrete State Spectrum
In this section we introduce a vacuum self-regulation mechanism. It controls cell distribution among the attractors.
We assume that during their evolution, charge trajectories converging to a given attractor do not disappear. They squeeze together during cell evolution, and their density  increases with elapsing time in inverse proportion to the uncertainty intervals
increases with elapsing time in inverse proportion to the uncertainty intervals
 (2.31)
(2.31)
where  is initial trajectory density, and uncertainty intervals
is initial trajectory density, and uncertainty intervals  after a few iteration steps can be described as
after a few iteration steps can be described as
 (2.32)
(2.32)
Being an open system, a vacuum cell, under influences of random external forces, occasionally jumps from one trajectory to another. We define vacuum temperature  as average frequency of these jumps
as average frequency of these jumps
 (2.33)
(2.33)
where  is the average dwell time at a single trajectory.
is the average dwell time at a single trajectory.
We assume also that a jumping cell has equal probabilities to “land” onto any trajectory.
With the above assumptions, the probability  to find a cell near attractor with energy
to find a cell near attractor with energy  is proportional to the average trajectory density
is proportional to the average trajectory density 
 (2.34)
(2.34)
By combining (2.33) and (2.34), and replacing , we obtain that in zone
, we obtain that in zone  at temperature
at temperature , partition function is
, partition function is
 (2.35)
(2.35)
where  is a normalization constant. Using (2.28) and (2.29), we can extent (2.35) to the other zones
is a normalization constant. Using (2.28) and (2.29), we can extent (2.35) to the other zones
 (2.36)
(2.36)
where  is the zone number, and
is the zone number, and  is the corresponding chemical potential.
is the corresponding chemical potential.
Distribution (2.36) is the equation of vacuum self-regulation. It provides the probability to find a cell near the corresponding attractor (energy). Its form remarkably resembles that of the Boltzmann factor. The major difference is that the latter normally provides distribution of free particles (positive energies), while partition function (2.36) controls distribution of bounded cells (negative energies).
Partition function (2.36) can be rewritten for cell distribution in B-space. Three examples of  are shown in Figure 15 for three different temperatures (dwell times
are shown in Figure 15 for three different temperatures (dwell times ). The curves are normalized as
). The curves are normalized as
 (2.37)
(2.37)
At very high temperatures, when cell dwell time at a single trajectory decreases down to the length of the iteration interval , the distributions acquires form of a continuous spectrum (blue and green curves in Figure 15). At lower temperatures, dwell time
, the distributions acquires form of a continuous spectrum (blue and green curves in Figure 15). At lower temperatures, dwell time  is large enough for a cell to make a few rotations before it jumps to another trajectory, the partition function acquires the shape of the Dirac δ-function (red curve), and the cell distribution spectrum becomes essentially discrete.
is large enough for a cell to make a few rotations before it jumps to another trajectory, the partition function acquires the shape of the Dirac δ-function (red curve), and the cell distribution spectrum becomes essentially discrete.
At normal temperatures, the vast majority of cells are located at the superattractors, and the superattractor characteristics acquire rank of physical constants. They are not absolute but may deviate from the superattractor values under action of external forces and temperature, i.e. these constants are “running”. (Recall the running coupling constants in the standard model, like electron charge).

Figure 15. Normalized cell distributions among attractors (B) at temperatures  , and
, and .
.
2.7. Chemical Potentials and Electroweak Mixing Angles
Let us return to very high temperatures, when cellular jumps among the trajectories become so frequent, that cells do not have enough time to complete even a single rotation. At these temperatures, cells lose their cellular features and dissolve in the background dust. The critical temperature of the cellular “melting” is
 (2.38)
(2.38)
where  is the cell rotation period.
is the cell rotation period.
If “melted” vacuum cools down to the temperatures below critical,  , cellular formation starts again. Vacuum can produce cells of different types that belong to different zones of stability. The number of cells of each type depends on the corresponding chemical potential
, cellular formation starts again. Vacuum can produce cells of different types that belong to different zones of stability. The number of cells of each type depends on the corresponding chemical potential .
.
Let us illustrate this on a simple example, when the resulting cell mixture consists only of two types of cells, Z1-cells and Z2-cells. According to our associations with the standard model, these cells represent respectfully electromagnetic interactions and weak nuclear interactions. We want to estimate their relative input, i.e. ratio , where
, where  and
and  are probabilities to find respectively Z1-cells and Z2-cells. In a two-component mixture, the total probability is
are probabilities to find respectively Z1-cells and Z2-cells. In a two-component mixture, the total probability is
 (2.39)
(2.39)
We can replace  and
and  with trigonometric functions of a single parameter, angle
with trigonometric functions of a single parameter, angle , which we call a mixing angle
, which we call a mixing angle
 (2.40)
(2.40)
Let us assume that at the melting point the temperature is fixed and is . Then, using (2.29) and (2.36), one can find that the
. Then, using (2.29) and (2.36), one can find that the  is also fixed and is
is also fixed and is
 (2.41)
(2.41)
Here ,
,  , and
, and  is the Feigenbaum delta.
is the Feigenbaum delta.
Using (2.40) and (2.41), one can find that the mixing angle  is also fixed and is
is also fixed and is
 (2.42)
(2.42)
Recall, that in the standard model, there exists a fixed number , which value is close to
, which value is close to , which also calls “mixing angle” (the Weinberg mixing angle or electroweak mixing angle), because it provides the relative input of electromagnetic (B0-boson) and weak (W0-boson) components in their mixture that calls photon.
, which also calls “mixing angle” (the Weinberg mixing angle or electroweak mixing angle), because it provides the relative input of electromagnetic (B0-boson) and weak (W0-boson) components in their mixture that calls photon.
The Weinberg mixing angle is an empirical constant. It is connected to the electromagnetic and weak interaction coupling constants  and
and  via Equation (recall (1.8))
via Equation (recall (1.8))
 (1.8)
(1.8)
By comparing (1.8) and (2.41), one can deduce that the coupling constants can be expressed via cellular chemical potentials
 (2.43)
(2.43)
This remarkable result suggests that the standard model coupling constants are thermodynamical characteristics of the corresponding fields.
Extending the model to three-component mixture is straightforward. Adding Z3-cells associated with the strong nuclear interactions, and including the strong coupling constant  in (2.43) gives us the proportion
in (2.43) gives us the proportion
 (2.44)
(2.44)
that is in agreement with(1.13)!
2.8. Anti-Cells
Charge conjugation is the only component of CPT-symmetry, which has an analog in the proposed model.
If in a given cell we invert directions of all radial flows, we obtain a complimentary charged cell. In the standard model, anti-particles can be represented as particles evolving backward in time. This possibility is inherited from Hamiltonian framework where time reversibility is one of the fundamental symmetries. In dissipative framework where the proposed model belongs to, we cannot revert time direction. This would convert all attractors into repellers and instead of asymptotic stability we obtain chaos. To invert intracellular flow directions without reversing time direction, we will use specific features of the even iteration function. To describe anti-cells, we will replace the original iteration function with its negative copy
 (2.45)
(2.45)
The inverted map (2.45) evolves in the same time direction. It possesses all features of the original map (2.5). One can find exactly the same special points at the same places, the same parameter values for  and
and , the same relations among them, the same levels of the asymptotic stability, the same probability distributions, etc. The only difference is that all intracellular flows are reverted and all charges have the opposite polarities.
, the same relations among them, the same levels of the asymptotic stability, the same probability distributions, etc. The only difference is that all intracellular flows are reverted and all charges have the opposite polarities.
The combined cell/anti-cell iteration function is
 (2.46)
(2.46)
where sign “+” stands for cells, and “?” for anti-cells.
The combined cell/anti-cell bifurcation diagram is shown in Figure 16. It comprises the original diagram that depicts fixed charge values for cells (shown in black) and the inverted diagram that depicts fixed charge values for anti-cells (shown in red).

Figure 16. Combined cell/anti-cell bifurcation diagram.
2.9. Discrete Cell Distribution Implies Quantized Charges
In section 2.6, we found that, for not extremely hot vacuum, cell distribution among the attractors is practically discrete. Almost all the cells are located near the superattractors, and superattractor characteristics become physical constants. The superattractor charges are among these special numbers. The charg- ed superattractor states are shown on the bifurcation diagram in Figure 17 by small color circles. Each charged state has its counterpart (the anti-state with the opposite charge polarity). Beside the charged superattractor states there exist neutral superattractor states. They are shown by white circles.

Figure 17. Quantized charges of the proposed model are shown by color circles.
In zone , which we associate with electromagnetic field, the only superattractor state is neutral,
, which we associate with electromagnetic field, the only superattractor state is neutral,  , and this is in agreement with electrically neutral photons.
, and this is in agreement with electrically neutral photons.
In zone , which we associate with weak nuclear field, there exist one neutral superattractor state and two complimentary charged states (purple circles). The charged states are in agreement with weak charge spectrum. Recall that in weak interactions, there exist neutral mediators,
, which we associate with weak nuclear field, there exist one neutral superattractor state and two complimentary charged states (purple circles). The charged states are in agreement with weak charge spectrum. Recall that in weak interactions, there exist neutral mediators,  boson, and charged mediators,
boson, and charged mediators,  bosons, carrying charges
bosons, carrying charges .
.
In zone , which we associate with strong nuclear field, there exist one neutral state and three pairs of complimentary charged states shown by red, green, and blue circles. The charged states are in agreement with the color charge spectrum carried by gluons, and we can associate red, green, and blue superattractor charges with color charges of chromodynamics.
, which we associate with strong nuclear field, there exist one neutral state and three pairs of complimentary charged states shown by red, green, and blue circles. The charged states are in agreement with the color charge spectrum carried by gluons, and we can associate red, green, and blue superattractor charges with color charges of chromodynamics.  neutral state has no analog in the standard model, but it does play an important role in explanation of flavor mixing amplitudes.
neutral state has no analog in the standard model, but it does play an important role in explanation of flavor mixing amplitudes.
One of the spinorial cell peculiarities is that their charged states are dynamical: if at one time instant, d-cell is at the neutral (white) state, the next instant, it is at the charged (purple) state. The charge pulsations can be leveled at some distance from the cells if we couple two d-cells and synchronize them out-of-phase. Then, when one of them is charged the other is neutral and vice versa. Another peculiarity is that in q-cells the charge states rotate always in the same order: …→ neutral → red → green → blue → neutral →…
Some important cellular quantum numbers we have encountered in the previous discussion are summarized in Table 2.
Table 2. Some important cellular quantum numbers.
3. Cellular Networks
In Section 2, we studied individual vacuum cells. In this part, we explore their networks.
3.1. Intercellular Dynamics. Synchronization between Two Cells
Synchronization is a ubiquitous phenomenon in self-organized systems [39] [40] [41] [42] . Since Huygens’s discovery in 1665, synchronization was observed in a number of active dissipative systems: coupled mechanical clocks, chemical reactions, electric and electronic circuits, animated cells, organs, and organisms, eco-systems, etc. Synchronization may happen between identical oscillators and between unlike devices, between couples and among components of a complex system, between oscillators producing near-sinusoidal waveforms and between chaos generators.
Vacuum cells are open systems and may couple to each other via dust exchanges as schematically shown in Figure 18. As self-sustained oscillators, they may synchronize. Synchronization is a dissipative process that occurs under repetition of delayed feedback loops. It is a phase transition that transforms independent vacuum cells into coherent networks. After synchronization, most of cells operate with one unified frequency (small amount of cells oscillate at different but commensurate frequencies). An important feature of synchronization is its universality: synchronized oscillators may differ among themselves by construction, geometry, topology, etc. This means that to synchronize, vacuum cells are not required to be identical. Below, we explore consequences of cell synchronization.

Figure 18. Cell coupling via flow exchanges.
We start with a simple system comprising two coupled phase oscillators, which prior to synchronization operated with different natural frequencies  and
and . Each oscillator imposes a periodical force onto the other oscillator. In a simplest case, the forces are sinusoidal. The strength of the force at each time instant depends on the current phase difference between the oscillators,
. Each oscillator imposes a periodical force onto the other oscillator. In a simplest case, the forces are sinusoidal. The strength of the force at each time instant depends on the current phase difference between the oscillators, . To describe phase dynamics, we use the Adler equation [39] :
. To describe phase dynamics, we use the Adler equation [39] :
 (3.1)
(3.1)
where  is initial frequency difference, and
is initial frequency difference, and  is a coupling parameter. We adapt this equation to discrete-time dynamics and transform it to the form
is a coupling parameter. We adapt this equation to discrete-time dynamics and transform it to the form
 (3.2)
(3.2)
where  is the phase difference immediately after
is the phase difference immediately after
 is the iteration time interval. To simplify notations, we set
is the iteration time interval. To simplify notations, we set , and obtain iterated map
, and obtain iterated map
 (3.3)
(3.3)
If the oscillators have different natural frequencies and there is no coupling between them, the phase difference  grows without limitation. When coupling is strong enough, the phase difference
grows without limitation. When coupling is strong enough, the phase difference  may converge to a fixed point
may converge to a fixed point . The phenomenon is known as phase entrainment or phase locking.
. The phenomenon is known as phase entrainment or phase locking.
Map (3.3) possesses asymptotic stability  if initial frequency difference
if initial frequency difference  (Figure 19 central part). Otherwise, the map is diverging
(Figure 19 central part). Otherwise, the map is diverging .
.

Figure 19. Phase differences  for map (3.3) after
for map (3.3) after  iteration steps.
iteration steps.
A few examples of converging  trajectories are shown in Figure 20. Independent on initial phase difference
trajectories are shown in Figure 20. Independent on initial phase difference , the trajectories converge to fixed values, which depends on the initial frequency difference
, the trajectories converge to fixed values, which depends on the initial frequency difference . Trajectories belonging to different
. Trajectories belonging to different  are shown in different colors.
are shown in different colors.
Phase-difference ( ) dynamics are similar to charge (
) dynamics are similar to charge ( ) dynamics, which we studied in Section 2. At the beginning of evolution,
) dynamics, which we studied in Section 2. At the beginning of evolution,  trajectories belonging to different
trajectories belonging to different  entangle and are resolved only after some elapsing time. The uncertainty intervals
entangle and are resolved only after some elapsing time. The uncertainty intervals  shrink with time exponentially (Figure 21). Their slopes
shrink with time exponentially (Figure 21). Their slopes  depend on the initial frequency difference
depend on the initial frequency difference  and coupling
and coupling . The highest convergence rate occurs at superattractor
. The highest convergence rate occurs at superattractor . If
. If  is fixed, the highest convergence rate occurs when
is fixed, the highest convergence rate occurs when .
.

Figure 20. Examples of phase difference  evolution. Trajectories with different
evolution. Trajectories with different  -values are shown by different colors.
-values are shown by different colors.

Figure 21. Examples of uncertainty interval  evolution.
evolution.
After synchronization, both oscillators operate at the same frequency. The frequency difference is always zero and  does not provide much information about oscillators. This function is “delegated” to a new order parameter, fixed points
does not provide much information about oscillators. This function is “delegated” to a new order parameter, fixed points 
 (3.4)
(3.4)
We define energy  similarly to how we define it for a single cell (Equation (2.25)), but instead of the cell period we use frequency
similarly to how we define it for a single cell (Equation (2.25)), but instead of the cell period we use frequency  which is established after synchronization
which is established after synchronization
 (3.5)
(3.5)
Function  has form of a potential well (Figure 22). The well depth depends on coupling
has form of a potential well (Figure 22). The well depth depends on coupling . For stronger couplings, the well deepens and its walls gradually acquire logarithmic shape (Figure 22(left)). For small couplings, the well bottom is smooth and its shape more resembles cosine function (Figure 22(right)).
. For stronger couplings, the well deepens and its walls gradually acquire logarithmic shape (Figure 22(left)). For small couplings, the well bottom is smooth and its shape more resembles cosine function (Figure 22(right)).


Figure 22.  for
for  (left) and
(left) and  (right).
(right).
To compare shapes of  and
and  wells, we need to bring their arguments to the same scale. We do this with a help of normalized distance to the attractor
wells, we need to bring their arguments to the same scale. We do this with a help of normalized distance to the attractor , which in this case we define as
, which in this case we define as
 (3.6)
(3.6)
Wells  and
and with reduced arguments are shown side by side in Figure 23. The calculated points (rhombs) fall on red curves describing logarithmic functions
with reduced arguments are shown side by side in Figure 23. The calculated points (rhombs) fall on red curves describing logarithmic functions  and
and  respectively. The data suggest that for all β-values
respectively. The data suggest that for all β-values
 (3.7)
(3.7)
This equality suggests that the energy (thus the asymptotic stability) of two strongly coupled and synchronized cells is equal to the doubled energy of a single cell, which is in accord with energy extensivity.


Figure 23. Single cell potential well  (left) and potential well for strongly coupled and synchronized couple
(left) and potential well for strongly coupled and synchronized couple  (right).
(right).
Synchronization process we considered above describes operation of self- sustained oscillators with periodical waveforms and can be applied to the circular cellular flows. Beside circular, cells possess a periodic radial flows. In fact, both types of flows involve the same dust particles, and being nonlinear, tend to synchronize. For example, converging evolution functions  in Figure 3.1 have oscillating components with period
in Figure 3.1 have oscillating components with period .
.
To illustrate how radial flows may synchronize, we can employ Wick’s rotation, . This transformation converts real exponents
. This transformation converts real exponents  of converging evolutions into imaginary exponents
of converging evolutions into imaginary exponents  of periodical processes
of periodical processes
 (3.8)
(3.8)
Here to avoid confusion, we use two different parameters for imaginary time and real time, .
.
The synchronization schema is shown in Figure 24.

Figure 24. Illustration of synchronization schema under Wick’s rotation.
If in the real-time state-space, two cells have close energies , in the imaginary-time state-space, they have close frequencies,
, in the imaginary-time state-space, they have close frequencies,  ,
,  ,
, . Then they synchronize
. Then they synchronize  at frequency
at frequency , which in the real-time state-space corresponds to the energy
, which in the real-time state-space corresponds to the energy . In summary, we obtain that cells synchronize their energies
. In summary, we obtain that cells synchronize their energies .
.
Not all synchronized cells may have the same frequency. Small amount of them that are at some distance from the superattractors, may synchronize at commensurate frequencies , where
, where  and
and  are integers [41] . It is important that even in this case, the cells are subjects to the phase entrainment and their phase differences can be used as network characteristic (order parameter). For simplicity, in the following discussions we assume that all cells synchronize at a single frequency.
are integers [41] . It is important that even in this case, the cells are subjects to the phase entrainment and their phase differences can be used as network characteristic (order parameter). For simplicity, in the following discussions we assume that all cells synchronize at a single frequency.
A special case is synchronization of spinorial cells. The spinorial cell full period is  and their phase differences can be as big as
and their phase differences can be as big as
 (3.9)
(3.9)
where  is a small deviation from integer number of
is a small deviation from integer number of . This happen when the synchronized cells are located at different attractor loops (Figure 25). We define two principally different synchronization modes: in-phase synchronization,
. This happen when the synchronized cells are located at different attractor loops (Figure 25). We define two principally different synchronization modes: in-phase synchronization,  , and out-of-phase synchronization,
, and out-of-phase synchronization,  .
.


Figure 25. Out-of-phase synchronized d-cells (left) and q-cells (right). Black points show cell locations at their attractors at some time instant.
At normal temperatures, most of the cells are located near the superattractors, operate at close frequencies, and .
.
3.2. Emergence of Global Symmetries. Phase Difference as Order Parameter
Before being synchronized, cells rotate each with its own pace and count their local time independently. After synchronization, they form coherent networks. They clock time with the same rate. Synchronization is a phase transition that produces a major symmetry: global time scale.
This symmetry provides a basis for unification of scales of other physical parameters. The list includes energy , temperature
, temperature , asymptotic stability
, asymptotic stability , critical temperature of cell dissociation/formation
, critical temperature of cell dissociation/formation , and chemical potentials
, and chemical potentials , which were defined as derivatives of time intervals.
, which were defined as derivatives of time intervals.
In coupled cell networks, phase entrainment creates a new order parameter? phase difference  between the cells. If we zoom out a synchronized cellular network, its granular (cellular) structure smoothes out, and the network acquires features of fiber bundle (Figure 26), where the base space is physical space and the cellular dynamics are elevated to the fibers.
between the cells. If we zoom out a synchronized cellular network, its granular (cellular) structure smoothes out, and the network acquires features of fiber bundle (Figure 26), where the base space is physical space and the cellular dynamics are elevated to the fibers.

Figure 26. Zoomed-out synchronized cellular network resembles fiber bundle.
Due to the phase entrainment, the fibers are inherently connected, and the local phase difference  between cells (now between fibers) acquires function of a field variable.
between cells (now between fibers) acquires function of a field variable.
Each fiber stems from its own cell and inherits the cell topology. Respectively, we can discriminate fibers with simple rotations from the spinorial fibers. Fiber bundle constructions emergent from ,
,  , or
, or  cells, resemble respectively electromagnetic, weak, or strong fields of the standard model.
cells, resemble respectively electromagnetic, weak, or strong fields of the standard model.
Spinorial cells can form diverse out-of-phase synchronization patterns, and toy particles (as we call them) constructed from the spinorial cells inherit symmetries of their synchronization patterns.
Synchronized cellular networks comprise cells of different geometry and sizes. It does not concern with distances between the cells. The networks are not crystal lattices and they do not provide space gauges. However, it is still possible to calibrate space intervals on a premise that we have had a universal time scale and, in addition, postulate that network excitations propagate through the network with a constant speed . Borrowing this postulate from relativity, we can define a standard space interval
. Borrowing this postulate from relativity, we can define a standard space interval  as
as
 (3.10)
(3.10)
where  is the standard time interval (cell rotation period).
is the standard time interval (cell rotation period).
This definition obviously connects space and time scales, but it does not establish sameness between space and time. Time remains to be irreversible and space-independent.
 -based time scale is not the only time scale that emerges in cellular networks. Later we will discuss dilated time resembling that of general relativity.
-based time scale is not the only time scale that emerges in cellular networks. Later we will discuss dilated time resembling that of general relativity.
There exists one more symmetry related to our approach that is worth mentioning. The important feature of iteration function (2.6) is that it has one and the only extremum. This feature is called unimodality. Because of this feature, the function belongs to the class of Feigenbaum universality. If one replaces this function with another function from this class, most of the emergent phenomena we encounter in this paper will be preserved. (S)he would find a similar bifurcation diagram, alike probability distributions, the same sets of discrete states, quantized charges, spinorial cells, the same values of chemical potentials, and so on. Even the charge conjugation symmetry can be recovered with a new function if it has a smooth extremum, which can be approximated by a quadratic parabola, by properly selecting coordinate system and its origin.
3.3. Quantum Distributions
In Section 3.1, we considered synchronization phenomenon called phase entrainment. Each of coupled cells in the network forces the other cells to change their energies toward its own value. After iterative “negotiations” the cells come to a common trade-off value. If we have more than two cells in the network and some of them have been already synchronized, the synchronized cells have progressive advantage. This nonlinearity creates a positive feedback: the more cells in the network have been synchronized at a given energy, the higher is the probability to get another cell at the same energy. Like in the case with iron filing described above, synchronization spontaneously break the original, almost continuous, cell distribution and create a discrete spectrum (Figure 27). Cell synchronization also alternates the Boltzmann-like cell distribution (2.36) that we encountered in the previous chapters.

Figure 27. Synchronization alternates cell distribution among the energies.
To formalize the effect of phase entrainment, let us consider a group of cells some of which have been synchronized at states with energy . Let the rest of the cells in this group have close but different energies. The synchronized cells force the latter to synchronize at the same energy. Let the initial probability to find a cell at state
. Let the rest of the cells in this group have close but different energies. The synchronized cells force the latter to synchronize at the same energy. Let the initial probability to find a cell at state  is 𝑤 and the resulting probability is
is 𝑤 and the resulting probability is . In simplest case, the probability to get a new cell at the same state is proportional to the number of cells at this state
. In simplest case, the probability to get a new cell at the same state is proportional to the number of cells at this state , where
, where  is the proportionality coefficient. The total number of the new cells is
is the proportionality coefficient. The total number of the new cells is , and the total number of all cells at state
, and the total number of all cells at state  after adding the new cells is
after adding the new cells is
 (3.11)
(3.11)
Signs “+” in (3.11) indicates that the synchronization leads to phase-entrain- ment (which is not the only possibility).
By solving Equation (3.11) for , we obtain that
, we obtain that
 (3.12)
(3.12)
and after substitution of Boltzmann-like factor , where
, where  is given by partition function (2.36), in (3.12), we obtain a new partition function
is given by partition function (2.36), in (3.12), we obtain a new partition function
 (3.13)
(3.13)
Remarkably, Equation (3.13) has Bose-Einstein distribution form. It is obtained without requirement of the identity of the participating cells!
Phase entrainment is not the only synchronization scenario. At the opposite pole, synchronization may quench oscillations [40] [45] [46] [47] [48] , the phenomenon known also as amplitude death or oscillation cessation. Quenching scenario depends on the details of involved oscillators, their coupling mechanisms, and other conditions. Applying to vacuum cells, we assume that quenching destroys cells as self-organized entities and they just dissolve into the dust background. We assume that quenching is also affected by a positive feedback. However, this time the cells compete not with their rivals, but with the noisy background. The more cells ceased to exist the stronger is the background forces destroying the remaining cells.
To describe cell distribution under quenching scenario, we use equations similar to (3.11) where we replace probability  with
with , coefficient
, coefficient  with
with , and sign “+” in parentheses with sign “−” to keep
, and sign “+” in parentheses with sign “−” to keep . After these replacements we obtain that
. After these replacements we obtain that
 (3.14)
(3.14)
Signs “−” in (3.14) indicates that the synchronization leads to the cell- quenching.
By solving (3.14) for , we obtain that
, we obtain that
 (3.15)
(3.15)
and after substitution of  in (3.15), we obtain that cell distribution for quenching scenario is
in (3.15), we obtain that cell distribution for quenching scenario is
 (3.16)
(3.16)
Distribution (3.16) has Fermi-Dirac distribution form.
We demonstrated that synchronization may transform the original distribution of independent cells into distributions typical for quantum-mechanical objects. Unlike quantum mechanics, where quantum statistics are intimately connected to the strict identity of particles, dissipative cells synchronize and form quantum distributions even if they are not copies of each other. This is a big relief from the enormous constraint. This is the power of synchronization, the power of asymptotic stability.
3.4. Synchronized Spinorial Cells as Particle Building Blocks
In this paper, we consider three types of cells belonging to zones Z1, Z2 and Z3. Their attractors are shown in Figure 28. Each attractor loop has its own color in the correspondence with the accepted charge-color code shown in Figure 17.
White loops represent quasi-neutral states/phases, purple loop represents ±e-charged phases, and red, green, and blue loops represent color-charged phases. Color-charge phases are ordered. They create a cyclic semi-group. The order for cells (anti-cells) is always the same and unidirectional: … → neutral → red → green → blue → neutral →… .

Figure 28. Attractor loop diagrams for Z1, Z2, and Z3 type cells.
With a few exceptions, we consider cellular networks comprising mostly of Z1-cells having single-loop attractors ( ). When consider complexes of
). When consider complexes of  and
and  spinorial cells, which have more complex attractor topology (
spinorial cells, which have more complex attractor topology ( ), we assume that they are immersed in
), we assume that they are immersed in  network as topological defects. Two, or more, directly coupled spinorial cells constitute a toy particle.
network as topological defects. Two, or more, directly coupled spinorial cells constitute a toy particle.
Each couple of synchronized cells carries a connecting link, that represent a special interest. We associate the connecting links with mediators of the corresponding fields. In case of synchronized spinorial cells, the connecting links inherit spinorial features from their host-cells and we call them spinorial links. Unlike regular links that occur between connected Z1-cells, the spinorial links are localized. They cannot travel across the network without their host-cells. Like extremely heavy W-bosons and confinement gluons, the spinorial links are doomed to be internal particle features.
To synthesize a toy particle, we are equipped with only four types of building blocks:  spinorial cells and anti-cells, and
spinorial cells and anti-cells, and  spinorial cells and anti-cells. Their combinations provide six different link types. This number is multiplied by the number of different spinorial-cell synchronization patterns (Figure 25). Different links may have different coupling strengths and carry different energies, that should diversify particle masses. However, these particle features are out of the scope of this paper.
spinorial cells and anti-cells. Their combinations provide six different link types. This number is multiplied by the number of different spinorial-cell synchronization patterns (Figure 25). Different links may have different coupling strengths and carry different energies, that should diversify particle masses. However, these particle features are out of the scope of this paper.
Examples of spinorial links are illustrated in Figures 29(a)-(c). Not all of the possible spinorial links have their analogs in the standard model. For example, a link between a neutral spinorial cell and a charged spinorial cell shown in Figure 29(d), or a link between d-cell and q-cell.




Figure 29. Spinorial cell links as localized bosons (shown by wavy lines): (a) charged weak boson; (b) neutral weak boson; (c) green-red gluon (d) white-red gluon.
Despite that the synchronized cell networks not necessary create a lattice, it is tempting to draw a parallel between the cell connecting links and solid-state phonons. In this respect, we would associate links between the in-phase synchronized cells with acoustic phonons, and links between the out-of-phase synchronized spinorial cells as optical phonons. Like their phonon counterparts, we expect that the “acoustic” links and “optical” links have different dispersions and different masses.
While building the toy-particles from the spinorial cells, we avoid in-phase synchronization between directly connected spinorial cells (light “acoustic” links) assuming that they are not stable enough.
Out-of-phase spinorial links carry at least one charge. Their charges rotate in synchrony with their host cells. Here is an example of “gluon”-link charge rotation in time



To avoid long phrases, we provide nicknames to the spinorial cells. They are listed in Table 3.
Table 3. Nicknames for spinorial cells.
In the paper, we use a number of illustrations/diagrams to better communicate the ideas. In the following discussions we will use one more type of diagrams, circular time-diagrams that linked to the other type diagrams as it is shown in Figure 30.
Upper line diagrams illustrates position on the bifurcation diagram (Figure 30(a)) and temporal behavior of Z2-cells: waveform (Figure 30(b)), loop-dia- gram (Figure 30(d)), and circular-time diagram (Figure 30(c)).
Bottom line diagrams illustrate Z3-cell dynamics.
Each circular time-diagram represents one full period  of a single cell. Attractor loops are represented by colored sectors according to the used above charge-color code. The upper sector (twelve o’clock position) shows current charge (state). The diagrams rotate in time, always counterclockwise. The cell rims are black in contrast to the anti-cell rims, which are red. In addition, d-on and d’-on diagrams expose the cell charge polarity marked by “−” and “+” signs.
of a single cell. Attractor loops are represented by colored sectors according to the used above charge-color code. The upper sector (twelve o’clock position) shows current charge (state). The diagrams rotate in time, always counterclockwise. The cell rims are black in contrast to the anti-cell rims, which are red. In addition, d-on and d’-on diagrams expose the cell charge polarity marked by “−” and “+” signs.

Figure 30. (a) Bifurcation diagrams; (b) Charge trajectories; (c) Circular time diagrams; d: Attractor loops.
Examples of circular time diagrams of in-phase and out-of-phase synchronized spinorial couples are shown in Figure 31. The formers have alike sectors at identical positions.

Figure 31. Examples of in-phase synchronized couples (left) and out-of-phase synchronized couples (right).
When building the toy particles, we use the following rules:
the stable spinorial links are links between out-of-phase synchronized couples;
d-cells/anti-cells are the only carriers of the electric charge ( );
);
d-cells/anti-cells are also the only carriers of  -and
-and  -flavors;
-flavors;
q-arcs are the only carriers of color charges and  -flavors. They are electrically neutral;
-flavors. They are electrically neutral;
in-phase synchronized q-arc is a carrier of unstable  -flavor and represents an intermediate (unstable) state;
-flavor and represents an intermediate (unstable) state;
toy leptons consist of coupled d-cells;
toy hadrons consists of coupled d-cells and q-arcs.
Flavor assignments illustrated by circular time diagrams are shown in Figure 32.

Figure 32. Flavor assignment diagram. Small arrows attached to
To match the set of quark flavors of the standard model, we mix q-cells/anti- cells carrying  -flavors and d-cells/anti-cells carrying
-flavors and d-cells/anti-cells carrying  -flavors and obtain the desired flavor set as direct (Cartesian) product
-flavors and obtain the desired flavor set as direct (Cartesian) product
 (3.17)
(3.17)
We avoid fractional electric charges by representing hadrons as toy lepton/q-arc mixtures.
Full set of q-arc flavors  with
with  placed properly between
placed properly between  and
and  forms a cyclic semi-group (Figure 33). Group generator
forms a cyclic semi-group (Figure 33). Group generator  produces unidirectional
produces unidirectional  -rotation of one of the two coupled circular time diagrams. Each
-rotation of one of the two coupled circular time diagrams. Each  -diagram-rotation corresponds to cell transition from one attractor loop to the next one, or
-diagram-rotation corresponds to cell transition from one attractor loop to the next one, or  -phase shift in the cell dynamics. The reverse rotations are prohibited because of time irreversibility. Instead, we rotate the complimentary diagram in the same direction. We use this symmetry (or anti-symmetry) when discussing flavor mixing matrix.
-phase shift in the cell dynamics. The reverse rotations are prohibited because of time irreversibility. Instead, we rotate the complimentary diagram in the same direction. We use this symmetry (or anti-symmetry) when discussing flavor mixing matrix.

Figure 33. Q-arc flavors  form a cyclic semi-group.
form a cyclic semi-group.
Beside different compositions and synchronization patterns, toy cells may possess different stereometry, architectures, and spinorial links. For example, cell permutations inside a particle may create a new particle(s).
Anti-particles are obtained from the corresponding particles by replacing all cells with their inversed counterparts, while preserving the original geometry and synchronization patterns.
4. Toy Particles
We associate small groups of synchronized and directly linked spinorial cells with toy particles. Toy particles are immersed into Z1-cell “electromagnetic” network. There exist four types of the toy particle building blocks: d-on, d’-on, q-on, and q’-on. The diversity of toy particles comes not only from different compositions but also because of their diverse synchronization patterns and architectures. Below, we consider a few examples of toy particles. They do not exhaust all possibilities but rather illustrate the power and capabilities of cellular dynamical networks.
4.1. Toy Leptons Spin
We build toy leptons exclusively from d-cells/anti-cells. Each directly coupled pair is synchronized out-of-phase. One possible arrangement of toy-leptons is shown by their circular time diagrams in Figure 34.

Figure 34. Toy lepton family. Electric charge pattern mimics pattern of real particles.
The charge pattern of toy leptons mimics the charge pattern of the standard model leptons. Like real leptons, the toy lepton family consists of three generations. Each generation has one positively charged particle, one negatively charged particle and one ( and
and ) or two (
) or two ( ) neutral particles (in Figure 34 only one τ-neutrino is shown).
) neutral particles (in Figure 34 only one τ-neutrino is shown).
Toy electron  consists of two out-of-phase synchronized d-ons (Figure 34 and Figure 35)). When one cell of the couple is in the charged phase, the other cell is electrically neutral, and vice versa. These charge oscillations create impression that the charge is moving in space from one cell to the other and backward (see Figures 35). However, the charge “motions” in space are not real, and their apparent “speed” may exceed the speed of light without confronting the relativity principles. The surrounding cells response to the charge pulsations by inducting currents
consists of two out-of-phase synchronized d-ons (Figure 34 and Figure 35)). When one cell of the couple is in the charged phase, the other cell is electrically neutral, and vice versa. These charge oscillations create impression that the charge is moving in space from one cell to the other and backward (see Figures 35). However, the charge “motions” in space are not real, and their apparent “speed” may exceed the speed of light without confronting the relativity principles. The surrounding cells response to the charge pulsations by inducting currents . Currents cannot go directly through d-ons. Therefore, the currents spontaneously break symmetry and create current loops. The current directions and orientation are established randomly through their recurrent interactions with d-ons. Their stability is fragile and their patterns can be easy alternated by perturbations. However, at normal conditions, the current producing oscillating charge (
. Currents cannot go directly through d-ons. Therefore, the currents spontaneously break symmetry and create current loops. The current directions and orientation are established randomly through their recurrent interactions with d-ons. Their stability is fragile and their patterns can be easy alternated by perturbations. However, at normal conditions, the current producing oscillating charge ( ) is rather a constant. The cell oscillation period (
) is rather a constant. The cell oscillation period ( ) is also a constant. Therefore, the induced current loops and the magnetic moments they create are also constants. We associate the phenomenon with the toy particle spin. It is obviously due to the spinorial features of d-ons (two-loop attractors) and out-of-phase synchronization.
) is also a constant. Therefore, the induced current loops and the magnetic moments they create are also constants. We associate the phenomenon with the toy particle spin. It is obviously due to the spinorial features of d-ons (two-loop attractors) and out-of-phase synchronization.

Figure 35. Diagram explaining origin of toy particle spin. Pulsating charges of out-of- phase synchronized d-ons (two left diagrams) induct current loops and magnetic moments in the surrounding network (two right diagrams). Purple and white circles repre- sent d-ons in charged and neutral states respectively. Grey circles represent surrounding electromagnetic cells.
Returning to Figure 28, toy positron,  , consists of two out-of-phase synchronized d’-ons. A neutral couple of d-on and d’-on plays the role of the electron neutrino,
, consists of two out-of-phase synchronized d’-ons. A neutral couple of d-on and d’-on plays the role of the electron neutrino, .
.
The second generation ( and
and ) is constructed by adding one d-on and one d’-on to each particle of the first generation. The additions do not change the charge pattern, but rather increase the particle energy (mass) and, most likely, affects its stability (lifetime).
) is constructed by adding one d-on and one d’-on to each particle of the first generation. The additions do not change the charge pattern, but rather increase the particle energy (mass) and, most likely, affects its stability (lifetime).
The third generation ( and
and ) is obtained the same way as the second generation, by adding two more complimentary charged d-cells to each particle of the second generation. Unlike two previous generations, the approach allows to obtain two different tau-neutrinos,
) is obtained the same way as the second generation, by adding two more complimentary charged d-cells to each particle of the second generation. Unlike two previous generations, the approach allows to obtain two different tau-neutrinos,  (only one is shown in Figure 34).
(only one is shown in Figure 34).
4.2. Toy Hadrons
We assemble toy hadrons as combinations of q-arcs and d-cells/anti-cells. Q- arcs provide  -flavors to hadrons, and d-cells/anti-cells provide
-flavors to hadrons, and d-cells/anti-cells provide  -flavors and electric charges. Direct product of
-flavors and electric charges. Direct product of  -flavors and
-flavors and  -flavors provides a set of six flavors resembling those of the standard model quark flavors.
-flavors provides a set of six flavors resembling those of the standard model quark flavors.
Toy mesons consist of one q-arc couple and two d-cells/anti-cells. Coupled circular time-diagrams of a group of toy mesons comprising  -flavored q-arcs are shown in the left column in Figure 36. The group consists of one positively charged particle carrying flavor
-flavored q-arcs are shown in the left column in Figure 36. The group consists of one positively charged particle carrying flavor , one negatively charged particle carrying flavor
, one negatively charged particle carrying flavor , and two neutral particles carrying two different flavors
, and two neutral particles carrying two different flavors . The latter differ by their architectures: in one of them, d-on is connected to q’-on and d’-on is connected to q-on, in the other, d-on is connected to q-on and d’-on is connected to q’-on.
. The latter differ by their architectures: in one of them, d-on is connected to q’-on and d’-on is connected to q-on, in the other, d-on is connected to q-on and d’-on is connected to q’-on.
A similar meson quartet comprising  -flavored q-arcs is shown in the right column in Figure 36.
-flavored q-arcs is shown in the right column in Figure 36.

Figure 36. Quartets of  - and
- and  -flavored toy mesons have the same electric charge patterns as quartets of real
-flavored toy mesons have the same electric charge patterns as quartets of real  - and
- and  -mesons that constitute eight-way octet shown in the diagram at right.
-mesons that constitute eight-way octet shown in the diagram at right.
The two toy-meson quartets have the same charge pattern as octet of real particles shown in “eight-way” diagram at the right.
In the same way we can assemble a meson quartet carrying  -flavored q-arcs.
-flavored q-arcs.
We can significantly extend the toy meson family by adding other geometries, like “stars”, “open chains”, and “tetrahedrons” shown in Figure 37.

Figure 37. Different toy mesons as spinorial cell “molecules”.
By adding one more q-arc to a toy meson we obtain a toy baryon. The baryons carry the same charge patterns but the number of different flavor combinations is significantly increased. Baryon flavors are direct products of

In Figure 38 we show an example of a toy baryon octet based on  - and
- and  -flavored q-arcs. The baryons have octahedron geometry. To prevent the diagram obstruction, links between d-ons and q-ons are not shown. The octet consists of two positively charged particles, two negatively charged particles, and four neutral particles. The toy baryon octet has a charge pattern similar to that of the real baryon octet comprising
-flavored q-arcs. The baryons have octahedron geometry. To prevent the diagram obstruction, links between d-ons and q-ons are not shown. The octet consists of two positively charged particles, two negatively charged particles, and four neutral particles. The toy baryon octet has a charge pattern similar to that of the real baryon octet comprising  and
and  quarks (Figure 38, the diagram at right). Two
quarks (Figure 38, the diagram at right). Two  -flavored particles (left column) are associated with proton and neutron.
-flavored particles (left column) are associated with proton and neutron.

Figure 38. Octet of  - and
- and  -flavored toy baryons has the same electric charge pattern as octet of real baryons consisting of u-, d-, and s-quarks (“eight-way” diagram at right).
-flavored toy baryons has the same electric charge pattern as octet of real baryons consisting of u-, d-, and s-quarks (“eight-way” diagram at right).
An example of “chemical reaction” between toy particles is shown in Figure 39. It describes exchange of d-on and d’-on between toy baryon and toy lepton accompanied by 2π-phase shift. In reality, neutrino should be replaced with anti-neutrino and moved to the right part of the equation. We write it in the presented form to simplify the picture.

Figure 39. Example of toy particle reaction mimicking neutron disintegration: .
.
4.3. Q-Arc Quartet and Flavor Mixing Matrix
The conventional six quark flavors constitute  flavor mixing matrix, which we briefly discussed in Section 1.3. We have observed that the matrix elements
flavor mixing matrix, which we briefly discussed in Section 1.3. We have observed that the matrix elements  can be approximated by the powers of the Feigenbaum delta
can be approximated by the powers of the Feigenbaum delta . The approximate CKM’-matrix is shown in Figure 40(left). We also found that
. The approximate CKM’-matrix is shown in Figure 40(left). We also found that  -matrix can be expanded to more symmetric
-matrix can be expanded to more symmetric  matrix, which is shown in Figure 40(right).
matrix, which is shown in Figure 40(right).

Figure 40. Approximate  -quark mixing matrix (left) and
-quark mixing matrix (left) and  -matrix q-arc mixing matrix that includes
-matrix q-arc mixing matrix that includes  flavor corresponding to in-phase synchronized q-arcs.
flavor corresponding to in-phase synchronized q-arcs.
Q-cells in q-arcs can synchronize in four different patterns. We associate these patterns with four q-arc flavors . They are shown in the
. They are shown in the  -matrix in the upper line and left column in the proper order. The matrix elements related to the “unstable” in-phase synchronized q-cells and carrying
-matrix in the upper line and left column in the proper order. The matrix elements related to the “unstable” in-phase synchronized q-cells and carrying  flavor are shown in pink. Remarkably, the pink cells accommodate all the Feigenbaum delta powers missing in
flavor are shown in pink. Remarkably, the pink cells accommodate all the Feigenbaum delta powers missing in  matrix. Moreover,
matrix. Moreover,  matrix elements are ordered! Their orders are shown by black and red arrows in Figure 41.
matrix elements are ordered! Their orders are shown by black and red arrows in Figure 41.

Figure 41. Matrix elements represent an ordered set of Feigenbaum delta powers.
Each arrow starts and ends at the same diagonal element. Moving from one matrix element to another along the arrow is accompanied by  -times reduction of the corresponding mixing amplitude
-times reduction of the corresponding mixing amplitude . According to q-arc synchronization schema, each step along the arrow corresponds to
. According to q-arc synchronization schema, each step along the arrow corresponds to  -rotation in q-cell dynamics, or in the other words, to a jump from one attractor loop to the next one (see Figure 42(central diagram)) or one-sector rotation of one of the two coupled circular diagrams (Figure 42(left and right diagrams)).
-rotation in q-cell dynamics, or in the other words, to a jump from one attractor loop to the next one (see Figure 42(central diagram)) or one-sector rotation of one of the two coupled circular diagrams (Figure 42(left and right diagrams)).

Figure 42. Examples of one-step shift along the red arrow (left) and three-step shift along the black arrow (right), and the corresponding jumps between the attractor loops (center).
All q-arc synchronization patterns and flavor changes covered by  matrix are shown in Figure 43 by means of coupled circular time diagrams.
matrix are shown in Figure 43 by means of coupled circular time diagrams.


Figure 43.  q-arc flavor changing matrix represented by coupled circular time-di- agrams. Each diagram couple is marked by the corresponding row/column number and the flavor name.
q-arc flavor changing matrix represented by coupled circular time-di- agrams. Each diagram couple is marked by the corresponding row/column number and the flavor name.
Moving farther, we describe transitions from one attractor loop to another (Figures 42(center)) as “tunnel” jumps (Figure 44) between the two synchronization states. The specificity of this “tunneling” is that it occurs not in the real space, but rather in the state-space. We can also say that this is tunneling in time.

Figure 44. “Tunneling in time”. Diagram describes the same processes as shown in Figure 42.
Using the analogy with the space tunneling, we estimate the probability  of loop-to-loop “tunnel” jumps. We replace tunnel distances with temporal intervals
of loop-to-loop “tunnel” jumps. We replace tunnel distances with temporal intervals
 (4.1)
(4.1)
where  is time required for a cell to complete a single loop rotation, which is also the iteration time interval,
is time required for a cell to complete a single loop rotation, which is also the iteration time interval,  is the number of steps along the arrows in Figure 41 diagram and also power in
is the number of steps along the arrows in Figure 41 diagram and also power in .
.
We assume that the tunnel barrier height  is equal to the q-arc internal energy
is equal to the q-arc internal energy , which is approximately doubled energy of a single q-cell or
, which is approximately doubled energy of a single q-cell or  (
( is chemical potential in zone
is chemical potential in zone )
)
 (4.2)
(4.2)
where  is q-cell rotation period.
is q-cell rotation period.
Now, we use the tunnel exponent to estimate the loop-to-loop jump probability which is
 (4.3)
(4.3)
After substitution of (4.1) and (4.2) into (4.3), we obtain that probabilities  are equal to the matrix elements
are equal to the matrix elements 
 (4.4)
(4.4)
Like all the phenomena we explore in this paper, q-arc flavor transformations are time irreversible. The direction of q-cell rotation is fixed and cannot be reversed, including the direction of the tunnel jumps. If we let cells to tunnel in both directions, the small forward three-step forward jumps probabilities,  , would be masked by much bigger one-step backward probabilities
, would be masked by much bigger one-step backward probabilities  and the mixing matrix elements would have different values and the matrix itself would have quite different appearance.
and the mixing matrix elements would have different values and the matrix itself would have quite different appearance.
4.4. “Dark Matter”
The list of electrically neutral toy particles we discussed in previous sections can be extended by adding other toy particles. Some examples are shown in Figure 45. The picture shows free q-arcs and based on them bigger complexes, and weakly and strongly interacting large cellular clusters. Being electrically neutral, they barely interact with electromagnetic field but carry energy (mass) and together with the previously considered neutral toy particles, can be associated with weakly and strongly interacting “dark” matter.

Figure 45. Examples of strongly and weakly interacting electrically neutral spinorial-cell assemblies.
5. “Relativistic” Phenomena and the Dark Zone
5.1. Time Dilation and Related Topics
We have seen that cell rotation period  can be used as a natural time unit in synchronized cellular networks. However, we did not provide any clue how to use it in practice. The problem is that with the existing technology and arsenal of instruments, we cannot trace the dust particle trajectories and measure time intervals linked to
can be used as a natural time unit in synchronized cellular networks. However, we did not provide any clue how to use it in practice. The problem is that with the existing technology and arsenal of instruments, we cannot trace the dust particle trajectories and measure time intervals linked to . Any imaginary clock would consist of at least two cells, and the clock rate would depend on how quickly the cells can establish their mutual dynamics. Normally, the speed of intercellular interactions is slower than intracellular relaxation, and is parametrized by parameters
. Any imaginary clock would consist of at least two cells, and the clock rate would depend on how quickly the cells can establish their mutual dynamics. Normally, the speed of intercellular interactions is slower than intracellular relaxation, and is parametrized by parameters ,
,  , or energy
, or energy . It is more convenient to have a measure of cell relaxation rate expressed as time intervals. We define cellular relaxation time
. It is more convenient to have a measure of cell relaxation rate expressed as time intervals. We define cellular relaxation time  as
as
 (5.1)
(5.1)
We assume that the clock rate is limited by the cellular relaxation time, and use  as a time unit for a “practical” scale.
as a time unit for a “practical” scale.
Unlike the cell rotation period , relaxation time
, relaxation time  is not network invariant. Different network regions may have different energies thus different τ-scale based clock rates. Using “practical” time unit
is not network invariant. Different network regions may have different energies thus different τ-scale based clock rates. Using “practical” time unit , we can define “practical” distance unit
, we can define “practical” distance unit  as
as
 (5.2)
(5.2)
where  is the speed of light.
is the speed of light.
By this definition, we connect space metric to the time metric. This connection is formal and does not established physical alikeness of space and time. A principal difference is that space is reversible while time is not. Formally, we can proceed even farther and convert Euclidian space-time into Minkowski’s space- time using Wick’s rotation, . In this case, definition (5.2) should be replaced with this one
. In this case, definition (5.2) should be replaced with this one
 (5.3)
(5.3)
Formally, Wick’s rotation would transform dissipative processes, described by the real time exponents, into periodical processes, described by imaginary exponents. i.e. the self-organized vacuum would be transformed into a conservation obeying medium, something resembling vacuum employed by the relativity theory or standard model of particle interactions.
With or without Wick’s rotation, “practical” time gauged by  resembles dilated time in general relativity. Drawing a parallel between the proposed model and general relativity, one can find that the flat space-time happens at superattractors, and the black holes emerge at bifurcations. Remarkably, the black hole emergence is accompanied by phase transitions from one type of fundamental interactions to another, starting from emergence of cellular structure (electromagnetic interactions) through emergence of weak, and strong fields. The initial state is lack of electromagnetic interactions and we call it “dark substance”. It locates in
resembles dilated time in general relativity. Drawing a parallel between the proposed model and general relativity, one can find that the flat space-time happens at superattractors, and the black holes emerge at bifurcations. Remarkably, the black hole emergence is accompanied by phase transitions from one type of fundamental interactions to another, starting from emergence of cellular structure (electromagnetic interactions) through emergence of weak, and strong fields. The initial state is lack of electromagnetic interactions and we call it “dark substance”. It locates in  or “the dark zone”.
or “the dark zone”.
5.2. The Dark Zone
In this section we briefly explore interval , which is located at the left of
, which is located at the left of  in the bifurcation diagram. This is vacuum state that gives birth to all fundamental interactions. We call it the dark zone. Another reason to give this name is that in
in the bifurcation diagram. This is vacuum state that gives birth to all fundamental interactions. We call it the dark zone. Another reason to give this name is that in , amplification parameter
, amplification parameter , and vacuum dust self- repelling is not balanced by its self-attraction. The vacuum dust progressively disperses when we approach the superattractor (
, and vacuum dust self- repelling is not balanced by its self-attraction. The vacuum dust progressively disperses when we approach the superattractor ( ), i.e. behave like under action of the dark energy.
), i.e. behave like under action of the dark energy.
The dark zone has some similarities with the other zones, but it also has a lot of differences.  -trajectories converge to a fixed point (Figure 46(left)), and this is the only fixed point,
-trajectories converge to a fixed point (Figure 46(left)), and this is the only fixed point, . The trajectories belonging to different
. The trajectories belonging to different  values always entangle with each other. They exponentially converge toward the only fixed point (Figure 46(right)). However, their slopes
values always entangle with each other. They exponentially converge toward the only fixed point (Figure 46(right)). However, their slopes  are different for different
are different for different  values. Formally, we can define energy
values. Formally, we can define energy  relaxation time
relaxation time , and delayed “practical time scale” using the same definitions as we used for the other zones.
, and delayed “practical time scale” using the same definitions as we used for the other zones.


Figure 46. Examples of evolution of radial flows  (left). Examples of evolution of uncertainty intervals
(left). Examples of evolution of uncertainty intervals  in zone
in zone  (right).
(right).
Using (2.19) we can define parameter  to find out that functions
to find out that functions  and
and  are practically the same as in the other zones (Figure 47).
are practically the same as in the other zones (Figure 47).

Figure 47.  and
and  in all semi-zones. Dark zone data are shown by blue circles.
in all semi-zones. Dark zone data are shown by blue circles.
The dark zone possess a strange stability. From the state-space point of view, this is a zone of asymptotic stability. Indeed, formally all trajectories asymptoti- cally converge toward the unique fixed point (which is also the superattractor). From the real physical space point of view, the vacuum dust unboundly disperse across the space. The closer the dust locates to the fixed point the stronger is the dispersion rate in space. In the dark zone, vacuum dust behaves like a chaotic system. The exponential expansion of the dust in space also resembles the Hubble law.
6. Concluding Remarks
In contrast to the existing tradition, we introduced a new dynamical model without any reference to Hamiltonian, Lagrangian, or variational principle. In the proposed model, the role of those is delegated to the iteration functions , which comprise all what we need to describe the vacuum cells evolution.
, which comprise all what we need to describe the vacuum cells evolution.
Curiously, in each zone,  can be written in a form resembling Lagrangian as difference between two functions,
can be written in a form resembling Lagrangian as difference between two functions,  and
and  (see (6.1)). The former is independent of variable
(see (6.1)). The former is independent of variable , represents external forces (through control parameter
, represents external forces (through control parameter ) and can be associated with the system potential energy. The other explicitly depends on
) and can be associated with the system potential energy. The other explicitly depends on , reflects the system reaction (through variable
, reflects the system reaction (through variable ), and can be associated with kinetic energy.
), and can be associated with kinetic energy.
 (6.1)
(6.1)
where

Newtonian, Lagrangian, Hamiltonian equations of motions in classical mechanics, Feynman’s pass integrals in quantum mechanics, Einstein-Hilbert general relativity equations can be “derived” with use of variational principles. The final results allow us to determine classical trajectories or most probable quantum states. Remarkably, we can formulate a “variational principle” for the vacuum cell dynamics. We have seen that the most probable locations for the vacuum cells in the state-space are super attractors. They always locate at the local extrema of the corresponding iteration function that is graphically illustrated in Figure 48. The pictures show graphical solutions of equations for super attractors ( ) in each of the three zones:
) in each of the three zones:
 (6.2)
(6.2)

Figure 48. Equation (6.2) graphical solutions for .
.
Left parts of equations,  , are represented by blue curves, and right parts,
, are represented by blue curves, and right parts,  , are represented by black and red diagonals. All stable solutions can be found at
, are represented by black and red diagonals. All stable solutions can be found at  local extrema
local extrema
 (6.3)
(6.3)
They are shown by small circles, white―for neutral states, and colored―for charged states.
Another conventional way to find location of stable states is to determine minimum of potential energy. Applying to the cell dynamics, we can use “potential functions”  defined in (6.1) to find the locations of super attractors. However, instead of function minima, we are looking for their roots (see Figure 49).
defined in (6.1) to find the locations of super attractors. However, instead of function minima, we are looking for their roots (see Figure 49).

Figure 49. Finding superattractors  as roots of potential functions,
as roots of potential functions, .
.
However, square of the potential functions form “potential wells” with minima at the superattractors (Figure 50).

Figure 50.  form potential wells with superattractor states at their bottoms.
form potential wells with superattractor states at their bottoms.
When we are talking about particle interactions such as weak or strong nuclear interactions, do we really believe that Nature is that intelligent that she invented the Dirac or Yang-Mills equations? Can we consider the standard model, or super-gravity, or strings, which operate with highly developed math as fundamental laws that control the universe?
Let us for a second assume that the toy particles are real, that we learned about their properties including explicit form of equations (6.1) from fancy experiments without any clue that they can be obtained by using a simple iteration process. Would we call (6.1) the fundamental Nature equations? Perhaps.
But we do know that the complexity and symmetries of the toy particles stem from dull iterations of a primitive function. We have observed quantum behavior, complex structures, symmetries and physical constants emergent on premises of system openness, dissipation, randomness, and competition rather than according to some intelligent design, in a striking similarity to Darwin’s evolution.
Acknowledgements
The paper is dedicated to the memory of my parents.
It would never be written if I did not meet my math teacher Isaak Rotfort and my university mentor Fernando Wilf. This project was born in connection with my daughter Julia’s discovery of curiously strange correlations in chaotic networks that resembles remote particle entanglement [49] . The proposed model went through a number of iterations under the influences of numerous papers, books, and Internet sources. The most relevant literature is listed below. I owe thanks to my colleagues Vladimir Litvinov and Lev Sadovnik, and my children Julia and Alexander for stimulating discussions, comments, and suggestions.
Cite this paper
Manasson, V.A. (2017) An Emergence of a Quantum World in a Self-Organized Vacuum―A Possible Scenario. Journal of Modern Physics, 8, 1330-1381. https://doi.org/10.4236/jmp.2017.88086
References
- 1. Hieb, M. (1995) Feigenbaum’s Constant and the Sommerfeld Fine-Structure Constant. Unpublished.
- 2. Goldfain, E. (2006) Chaos, Solitons & Fractals, 30, 324-331.
https://doi.org/10.1016/j.chaos.2006.01.117 - 3. Manasson, V.A. (2006) Self-Interacting Electron as a Nonlinear Dynamical System.
https://arxiv.org/abs/nlin/0609043 [nlin.CD] - 4. Manasson, V. A. (2008) Are Particles Self-Organized Systems?
https://arxiv.org/abs/0803.3300 [physics.gen-ph] - 5. Feigenbaum, M.J. (1978) Journal of Statistical Physics, 19, 25-52.
https://doi.org/10.1007/BF01020332 - 6. Feigenbaum, M.J. (1979) Journal of Statistical Physics, 21, 669-706.
https://doi.org/10.1007/BF01107909 - 7. Coullet, P. and Tresser, C. (1978) Le Journal de Physique Colloques, 539C, 5-25; C.R. Acad. Sci. Paris, 287A, 577-580.
- 8. Grossmann, S. and Thomae, S. (1977) Z. Naturforsh, 32a, 1353-1363.
- 9. Collet, P. and Eckmann, J.-P. (1980) Iterated Maps on the Interval as Dynamical Systems. Birkhauser, Basel.
- 10. Cvitanović, P. (1984) Universality in Chaos. Adam Hilger, Ltd., Bristol.
- 11. Cvitanović, P. (1984) Acta Physica Polonica, A65, 203-239.
- 12. Hilborn, R.C. (1994) Chaos and Nonlinear Dynamics: An Introduction for Scientists and Engineers. Oxford University Press, New York, Oxford.
- 13. Strogatz, S.H. (1994) Nonlinear Dynamics and Chaos: With Applications to Physics, Biology, Chemistry, and Engineering. Perseus Books Publishing, LLC, Westview.
- 14. Sfondrini, A. (2013) Introduction to Universality and Renormalization Group Techniques. Proceedings of Science Pos (Modave VIII), 005, Arxiv: 1210.2262v3 [Hep-Th] 25 Jun 2013.
- 15. Griffiths, D.J. (1987) Introduction to Elementary Particles. John Willey & Sons, Hoboken.
https://doi.org/10.1002/9783527618460 - 16. Physical Constants (2013) Particle Data Group Review.
http://pdg.lbl.gov/2017/reviews/rpp2016-rev-phys-constants.pdf - 17. The CKM Quark-Mixing Matrix (2014) Particle Data Group Review.
http://pdg.lbl.gov/2014/reviews/rpp2014-rev-ckm-matrix.pdf - 18. Benettin, G., Cercignani, C. Galgani, L. and Giorgilli, A. (1980) Lettere al NuovoCimento, 28, 1-4.
https://doi.org/10.1007/BF02776065 - 19. Greene, J.M., Mackay, R.S., Vivaldi, F. and Feigenbaum, M.J. (1981) Physica D: Nonlinear Phenomena, 3, 468-486.
https://doi.org/10.1016/0167-2789(81)90034-8 - 20. Bountis, T. (1981) Physica D: Nonlinear Phenomena, 3, 577-589.
https://doi.org/10.1016/0167-2789(81)90041-5 - 21. Widom, M. and Kadanoff, L. (1982) Physica D: Nonlinear Phenomena, 5, 287-292.
- 22. Mackay, R.S. (1993) Renormalisation in Area-Preserving Maps. World Scientific, Singapore.
https://doi.org/10.1142/2001 - 23. Turing, A.M. (1952) Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 237, 37-72. https://doi.org/10.1098/rstb.1952.0012
- 24. Prigogine, I. and Nicolis, G. (1977) Self-Organization in Non-Equilibrium Systems. Wiley, New York.
- 25. Haken, H. (1983) Advanced Synergetics: Instability Hierarchies of Self-Organizing Systems and Devices (Springer Series in Synergetics). Springer-Verlag, Berlin Heidelberg.
- 26. Prigogine, I. and Stengers, I. (1984) Order Out of Chaos. Bantam, New York
- 27. Nicolis, G. and Prigogine, I. (1989) Exploring Complexity. W. H. Freeman and Co., New York.
- 28. Wesfreid, J.E. (2006) Scientific Biography of Henri Bénard (1874-1939). In: Mutabazi, I., Wesfreid, J.E. and Guyon, E., Eds., Dynamics of Spatio-Temporal Cellular Structures, Springer, New York, 9-37. (See the List of Bénard’s Papers and Description of His Experiments)
- 29. Maurer, J. and Libchaber, A. (1979) Journal De Physique Lettres, 40, 419-423.
https://doi.org/10.1051/jphyslet:019790040016041900 - 30. Feigenbaum, M.J. (1979) Physics Letters A, 74, 375-378.
https://doi.org/10.1016/0375-9601(79)90227-5 - 31. Libchaber, A. and Maurer, J. (1980) Une Experience De Journal De Physique Colloques, 41, C3-51-C3-56.
- 32. Libchaber, A., Laroche, C. and Fauve, S. (1982) Period Doubling Cascade in Mercury. A Quantitative Measurement. Journal De Physique Lettres, 43, 211-216.
https://doi.org/10.1051/jphyslet:01982004307021100 - 33. Yahata, H. (1983) Progress of Theoretical Physics, 69, 1802-1805.
https://doi.org/10.1143/PTP.69.1802 - 34. Winfree, A.T. (1987) When Time Breaks Down. Princeton University Press, Princeton.
- 35. Walgraef, D. (1997) Spatio-Temporal Pattern Formation: With Examples from Physics, Chemistry, and Material Science. Springer-Verlag, New York.
https://doi.org/10.1007/978-1-4612-1850-0 - 36. Camazine, S., Deneubourg, J.-L., Franks, N.R., Sneyd, J., Theraulaz, G. and Bonabeau, E. (2001) Self-Organization in Biological Systems. Princeton University Press, Princeton and Oxford.
- 37. Schöll, E. (2001) Nonlinear Spatio-Temporal Dynamics and Chaos in Semiconductors. Cambridge University Press, Cambridge.
https://doi.org/10.1017/CBO9780511524615 - 38. Solé, R.V. and Bascompte, J. (2006) Self-Organization in Complex Ecosystems. Princeton University Press, Princeton and Oxford.
- 39. Adler, R. (1946) Proceedings of the IRE, 34, 351-357; Reprinted in Proceedings of the IEEE, 61, 1380-1385 (1973).
https://doi.org/10.1109/PROC.1973.9292 - 40. Strogatz, S.H. (2000) From Kuramoto to Crawford: Exploring the Onset of Synchronization in Populations of Coupled Oscillators. Physica D: Nonlinear Phenomena, 143, 1-20.
https://doi.org/10.1016/s0167-2789(00)00094-4 - 41. Pikovsky, A., Rosenblum, M. and Kurths, J. (2001) Synchronization. A Universal Concept in Nonlinear Sciences. Cambridge University Press, Cambridge.
https://doi.org/10.1017/CBO9780511755743 - 42. Balanov, A., Janson, N., Postnov, D. and Sosnovtseva, O. (2009) Synchronization: From Simple to Complex. Springer.
- 43. Dörfler, F. and Bullo, F. (2014) Automatica, 50, 1539-1564.
https://doi.org/10.1016/j.automatica.2014.04.012 - 44. Penrose, R. (2004) The Road to Reality: A Complete Guide to the Laws of the Universe. Vintage Books, New York.
- 45. Yamaguchi, Y. and Shimizu, H. (1984) Physica D: Nonlinear Phenomena, 11, 212-226.
- 46. Bar-Eli, K. (1985) Physica D: Nonlinear Phenomena, 14, 242-252.
https://doi.org/10.1016/0167-2789(85)90182-4 - 47. Mirollo, R.E. and Strogatz, S.H. (1990) Journal of Statistical Physics, 60, 245-262.
https://doi.org/10.1007/BF01013676 - 48. Koseska, A., Volkov, E. and Kurths, J. (2013) Physics Reports, 531, 173-199.
https://doi.org/10.1016/j.physrep.2013.06.001 - 49. Manasson, J. and Manasson, V.A. (2016) Strange Correlations between Remote Nodes in Networks Comprising Chaotic Links.
https://arxiv.org/abs/1511.02772 [nlin.CD]