Critical Nitrogen Dilution Curve in Processing Potato Cultivars ()
Received 27 October 2015; accepted 8 December 2015; published 11 December 2015


1. Introduction
Potato (Solanum tuberosum L.) crop frequently requires great application rates of nitrogen (N) fertilizer to achieve high tuber yield and quality. An adequate determination of the N fertilizer rate to be applied is important from both economic and environmental points of view [1] [2] . In potato, insufficient N leads to reduced yields, whereas excessive N leads to reduced yields and/or quality [3] . However, the optimal N fertilizer rate can vary widely among sites and among years. This variation results from the variation in both the N demand and the soil N supply. As a result, the development of tools to precisely predict the N requirement can optimize tuber yield and quality, while minimizing the risk of N losses to the environment [1] [4] .
Plant-based diagnostic tests that assess potato N sufficiency can be used to guide in-season N fertilizer management. The diagnosis of the plant N status during the growing season consists in the early detection of plant N deficiencies in order to determine the necessity of additional N fertilizer applications. Another application of plant-based diagnostic methods of N deficiency is diagnosis after harvest, aimed to detect limiting factors for crops within experimental sites or production fields. Plant-based diagnostic methods of N deficiency can be based on the determination of a critical N dilution curve [1] [4] . This function allows defining the critical N concentration (Nc) which is the minimum N concentration required for a maximum growth. The concept of a critical N dilution curve based on whole-plant N concentration studies was developed by Lemaire and Salette [5] and can be described empirically by a negative power function (Equation (1)) [6] [7] .
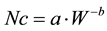 (1)
(1)
where W is the total biomass expressed in Mg∙ha−1, Nc is the critical N concentration in total biomass expressed in g 100 g−1, and a and b are fitting parameters. Coefficient a represents the N concentration when total biomass is at £1 Mg∙ha−1. The dimensionless coefficient b is referred as the “dilution coefficient”, which describes the decrease in N concentration caused by increasing total biomass. This power function can be used when total biomass ranges between 1 and 12 - 15 Mg∙ha−1 [6] [8] . For total biomass values lower than 1 Mg∙ha−1, Nc takes a constant value due to the small decrease in Nc with increasing total biomass [8] and the lack of competition for light of isolated plants [9] . On the other hand, the upper W limit in the function is defined by the maximum biomass that a crop can produce [6] [8] [10] .
The N concentration in individual plants within a dense canopy declines as the plant grows, even when there is an ample supply of N. This phenomenon has been usually interpreted as a result of plant ageing and plant phenology [11] . For the potato crop, the value of coefficients a and b is estimated using the combined biomass of shoots and tubers, and the N concentration of this total biomass. In potato, critical N dilution curves were first proposed in Scotland and the Netherlands by Greenwood et al. [6] , who reported values of 5.36 and −0.46 for a and b, respectively. Later, in France, Duchenne et al. [12] proposed values of 5.21 and −0.56 for a and b, respectively, and did not determine differences among different cultivars in critical N dilution curves. In a previous study where we determined the critical N dilution curve for the cultivar Innovator [13] , the estimated values for coefficients a and b were 5.30 and −0.42, respectively. These coefficients were similar to those reported by Greenwood et al. [6] and Duchenne et al. [12] . In contrast, in Canada, Belanger et al. [14] reported different coefficients of critical N dilution curves in two potato cultivars (Shepody and Russet Burbank). These cultivars had similar b coefficient but a different a coefficient. Shepody had a greater a coefficient (5.07) than Russet Burbank (4.57). These authors argued that this difference between cultivars was caused by differences in plant density. The cultivar with greater plant density had smaller plants with greater N concentration than the cultivars with low plant density. The Nc can be used to determine the plants N requirements and can be used to predict the N nutritional status of potato crop. Lemaire and Gastal [7] proposed an N nutrition index (NNI) to quantify the intensity of both N deficiency and luxury consumption by crops. The NNI for a given accumulated biomass can be estimated by the ratio between the biomass N concentration and the Nc. Values of NNI ≥ 1.0 indicate N sufficiency, whereas values of NNI lower than 1.0 indicate N deficiency. The Nc and the derived NNI are considered a reference tool for assessing crop N nutrition.
Potato cultivars differ in their biomass and N partitioning to leaves and tubers during the growing season. Kleinkopf et al. [15] observed that cultivars show differences in the proportions of N and dry matter accumulated in their different organs. Cultivars with shorter growing seasons have higher leaf growth rates at the initial stages of the growing season, higher harvest index (HI) and N harvest index (NHI) [16] , and lower N concentration in total biomass [17] than cultivars with longer growing seasons. Zebarth et al. [18] demonstrated that potato cultivars with different growth cycles have differences in biomass; N concentration and N use efficiency. Long-season cultivars produce more stem and leaves than short-season cultivars [19] [20] . This difference in growth patterns among potato cultivars could affect biomass N concentration. Thus, we hypothesize that morphologically different potato cultivars have different critical N dilution curves. These differences among cultivars may result in different nutritional N status. The results of this research could be used to improve management strategies for N fertilization because the critical N dilution curve would help to improve prediction of N demand during growing season in different potato cultivars. Our objectives were: a) to determine critical N dilution curves for specific potato cultivars; b) to compare these curves with reference curves determined by Greenwood et al. [6] , Duchenne et al. [12] and Bélanger et al. [14] ; c) to assess the possibility of using the critical N dilution curve to estimate the N status of the crop.
2. Materials and Methods
A field experiment was conducted during the 2003/4, 2004/5, 2005/6 and 2006/7 growing seasons at the McCain Argentina experimental field site, located in Balcarce (37˚45'S; 58˚18'W, 130 m a.s.l.; 870 mm mean annual rainfall; 13.7˚C mean annual temperature), Buenos Aires province, Argentina. The soils were Typic Argiudoll [21] Mar del Plata series, a particle size distribution of 240 g∙kg−1 clay, 330 g∙kg−1 silt, and 430 g∙kg−1 sand, and a minimum effective depth of 1.5 m. Soil organic matter (0 - 20 cm) was determined by the method of Walkley and Black [22] , pH (0 - 20 cm) with a glass electrode in a suspension of 1:2.5 soil/water ratio, available P (0 - 20 cm) by the method of Bray and Kurtz [23] , and 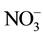 -N (0 - 40 cm) by the method of Bremner [24] and Keeney and Nelson [25] . The site characteristics for the 2003/4, 2004/5, 2005/6 and 2006/7 growing seasons were: soil organic matter (0 - 20 cm): 46, 46, 52 and 42 g∙kg−1; pH (0 - 20 cm): 5.7, 6.1, 6.2 and 6.0; P Bray-1 (0 - 20 cm): 21.1, 17.6, 19.3 and 30.2 mg∙kg−1; and NO3-N (0-40 cm): 52.1, 28.1, 12.9 and 25.7 kg∙ha−1, respectively. The previous crops were pasture (Bromus catharticus, Lolium perenne and Trifolium repens) (2004/05 and 2005/06 seasons) or soybean (Glycine max L) (2003/04 and 2006/07 seasons). Before planting, the soil was tilled using a disk harrow, a chisel and a vibrocultivator. Planting was carried out with a two-row planter with 0.85 m row spacing and an average seed density of 5 seed pieces per row meter. The planting density was 59,000 seeds ha−1.
-N (0 - 40 cm) by the method of Bremner [24] and Keeney and Nelson [25] . The site characteristics for the 2003/4, 2004/5, 2005/6 and 2006/7 growing seasons were: soil organic matter (0 - 20 cm): 46, 46, 52 and 42 g∙kg−1; pH (0 - 20 cm): 5.7, 6.1, 6.2 and 6.0; P Bray-1 (0 - 20 cm): 21.1, 17.6, 19.3 and 30.2 mg∙kg−1; and NO3-N (0-40 cm): 52.1, 28.1, 12.9 and 25.7 kg∙ha−1, respectively. The previous crops were pasture (Bromus catharticus, Lolium perenne and Trifolium repens) (2004/05 and 2005/06 seasons) or soybean (Glycine max L) (2003/04 and 2006/07 seasons). Before planting, the soil was tilled using a disk harrow, a chisel and a vibrocultivator. Planting was carried out with a two-row planter with 0.85 m row spacing and an average seed density of 5 seed pieces per row meter. The planting density was 59,000 seeds ha−1.
In the southeast Pampas region, processing potato cultivars of European and American origin have been introduced in recent years. In the present study, five cultivars with different morphological and agronomic traits were evaluated: Innovator (mid-season, 120 - 130 days from planting to crop maturity; determinate growth type), Gem Russet (late, 130 days, indeterminate growth type) [26] [27] , Umatilla Russet (medium late, 130 days; indeterminate growth type) [28] , Bannock Russet (late to very late, 140 days; indeterminate growth type) [27] [29] and Markies Russet (late to very late, 140 days; indeterminate growth type) [30] . In 2003/04, field trials were conducted with the potato cultivars Innovator, Gem Russet and Bannock Russet. In 2004/05 and 2005/6, field trials were conducted with the cultivars Innovator, Gem Russet, Umatilla Russet, Bannock Russet and Markies Russet. In 2006/7, field trials were conducted with the cultivars Innovator, Gem Russet and Umatilla Russet.
In each growing season, the cultivars were planted in a randomized complete block design with three replications and four N rates (Table 1). The N rate to each treatment was adjusted to soil test done before planting each season. The amount of N applied in treatments N0 and N1 represent fields with N deficiency. The amount of N applied in N2 was similar to the rate used by the farmers in the region. The amount of N applied in N3 exceeded the rate used by the farmers. The combination of fertilizer N rates allowed a wide range of N availability throughout the crop cycle. Fertilizer N rate for all growing seasons and cultivars were split at planting, hilling and bulking tuber. There is evidence that split applications are effective in increasing yield and N use efficiency in potato [27] [31] [32] . Each experimental unit had 12 rows and 14 m length. Planting was carried out on October 24th 2003, October 4th 2004, October 20th 2005 and November 5th 2006. The N source was granulated urea (NH2)2CO, which is generally used by local farmers, due to its low cost per N unit. The urea was surface broadcasted and then incorporated with 10 mm water irrigation. To ensure that P did not limit crop growth, triple super phosphate (NPK: 0-20-0) was applied at rates of 250 (2003/4, 2004/5 and 2005/6 seasons) and 180 (2006/7 season) kg∙ha−1. The soils of the field experiments have high content of K (> 400 ppm of extractable K) and K fertilization not used. Before canopy closure, hilling was also performed. Weeds, pests and fungal diseases were chemically controlled. Mechanical and chemical methods were used for weed control. The products used for weed control were Metaloclor (3 L∙ha−1) and Metribuzin (0.8 L∙ha−1). The control of diseases that normally appear in the potato crop, as Alternaria solani and Phytophthora infestans, was controlled with Mancozeb (at a dose a dose of 2.5 kg∙ha−1 and twelve applications are performed during the growing seasons) and with Chlorothalonil (at a dose of 1 L∙ha−1 and two applications were made during the growing season).
![]()
Table 1. Nitrogen rate and time of application in each treatment (N0, N1, N2 and N3) and growing season (2003/4, 2004/5, 2005/6, and 2006/7).
Irrigation was started at about 40 - 45 days after planting and continued until the end of the cycle. Water application was based on soil water potential assessed with Watermark sensors. Irrigation was initiated when the soil moisture was about 65 % of the water available in the soil profile. Irrigation was carried out with a forward advance sprinkler system Reinke Electrogator II (Reinke Deshler-Nebraska USA), starting on November 11th in 2003, November 27th in 2004, December 7th in 2005 and November 20th in 2006. The volume of water received from rainfall and irrigation was 610, 404, 514 and 756 mm in 2003, 2004, 2005 and 2006, respectively.
Shoot biomass and tuber biomass were measured at different times during each growing season, starting around 40 days after planting (DAP) until senescence, harvesting plants from 1 m of one row in each plot. Sampling dates were selected in order to have information from different stages of the crop cycle: vegetative growth (from emergence until 50 DAP), tuber initiation (50 - 60 DAP) and tuber bulking (60 DAP until physiological maturity) [33] . Shoot biomass and tubers were weighed fresh and subsamples (200 g each) were collected for determination of dry matter (DM) concentration and laboratory analyses. Subsamples were dried at 60˚C in a forced air draft oven for 7 d, ground to pass through a 1-mm sieve in a Wiley mill and stored at room temperature before laboratory analyses. The N concentration in shoots and tubers was determined using TruSpec CN analyzer [34] . Total biomass was calculated by adding shoot and tuber dry weight and was expressed in Mg DM ha−1. Total biomass N concentration (g 100 g−1) was calculated by adding the N content of shoots and tubers and dividing the obtained value by the total biomass. At harvest (March 11th 2004, February 22nd 2005, March 13th 2006 and March 20th 2007, respectively), 1 m of the two central furrows of each plot was harvested by hand to assess total tuber yield in fresh weight. A fraction of the samples was weighed and a sub-sample was dried and milled (0.84 mm mesh) to determine the dry matter content and N concentration [34] .
Data Analysis
The critical N dilution curve for each cultivar was determined using Equation (1), following the procedure proposed by Greenwood et al. [6] . The critical N dilution curve requires the identification of data points for which N does not limit biomass growth or is not in excess. Total biomass and N concentration for each cultivar, year and sampling date were subjected to analyses of variance (ANOVA) using the PROC GLM procedure [35] . Sampling dates were not used in determining the critical N dilution curves if the analysis of variance indicated no significant (p > 0.05) differences among N rate. For the remaining sampling dates, treatments were classified using the Fisher’s protected least significant difference (LSD) test. The highest total biomass obtained with any level of fertilizer and the corresponding N concentration were identified for each year and each sampling date and included in the database. When the highest total biomass was obtained in two or more levels of fertilizer, the data obtained with the lowest level were selected [6] [14] . These data points were used to determine the relationship between N concentration and total biomass using an allometric function (Equation (1)). When total biomass was below 1 Mg∙ha−1, we assumed that Nc took a constant value. The Nc for each cultivar was obtained using the SAS NLIN procedure [35] . To determine the Nc in Gem Russet, data from four growing seasons (2003/4, 2004/5, 2005/6 and 2006/7) were used. To determine the Nc in Umatilla Russet and Bannock Russet, data from three growing seasons (2004/5, 2005/6 and 2006/7; and 2003/4, 2004/5 and 2005/6 respectively) were used. To determine the Nc in Markies Russet data from two growing seasons (2004/5 and 2005/6) were used. We have previously determined the Nc for Innovator [13] , and this information was used for comparison among cultivars. The total biomass and N concentration were log-transformed to compare our fitted Nc and the reference Nc [6] [12] [14] . The slopes and intercepts of these linear regressions were analyzed using dummy variables [36] .
Differences in harvest index (HI) and N harvest index (NHI) among cultivars were evaluated using data from the 2004/5 and 2005/6 growing seasons, because the field trials with the five cultivars were conducted. The HI was calculated as the ratio between tuber biomass and total biomass for each N treatment and cultivar, while NHI was calculated as the ratio between tuber N and total N accumulated in each N treatment and cultivar. Treatment effects were evaluated by variance analysis using a PROC MIXED procedure included in the Statistical Analysis System program protocols [35] . The cultivars and N rate were considered as fixed effects and replications were considered as random effect. Then, a multiple comparison of means test (p < 0.05) was conducted with a Fisher’s protected least significant difference (LSD). The relationship of HI and NHI with the length of the period between planting and maturity was determined using linear regression analysis using PROC REG procedure. Additionally, the correlation between coefficient b, HI and NHI was determined.
Potato yield response to applied N fertilizer was determined for each cultivar by using PROC MIXED procedure (p < 0.05). The N rate was considered as fixed effect and replications were considered as random effect. The PROC NLIN procedure was then used to fit regression models for each cultivar and growing season identified as responsive to N rate. The quadratic regression model was used to predict the optimum N rate (ONR) for each cultivars and growing season. Nitrogen rate differential from the optimum N rate (dONR) was calculated as the ONR minus the applied N rates within each cultivar and growing season. Relative yield (RY) was calculated as the ratio between the tuber yield obtained at a given N rate and the highest tuber yield among all N application rates for each cultivar and growing season. The N nutrition index (NNI) was calculated at each sampling date and for each cultivar as the ratio between the measured N concentration of the whole plant and the predicted critical N concentration for the same total biomass [14] . The RY was expressed as a function of the NNI during tuber bulking (90 - 100 DAP) for each cultivar, and the quadratic regression model was fitted using the SAS NLIN procedure [35] . Similar relationship was determined between RY and dONR. To compare cultivars, the test of parallelism and coincidence was carried using a regression model containing dummy variables for the quadratic equation. Subsequently, each group of cultivars was fitted into a quadratic-plateau regression model using the SAS NLIN procedure [35] .
3. Results and Discussion
3.1. Critical N Dilution Curve in Potato Cultivars
Nitrogen concentration decreased as total biomass increased in all of the studied cultivars (Figure 1), coinciding with the results determined by Lemaire and Gastal [7] . This decline in N concentration with time, or increasing biomass, is attributed to a decrease in the fraction of total plant N associated with photosynthesis in relation to a concomitant increase in the N fraction of structural and storage constituents [7] . Innovator and Gem Russet had a lower plant N concentration than the other cultivars for a same level of total biomass. The Nc presented a constant value when total biomass values was lower than 1 Mg∙ha−1 [9] . The upper limit of total biomass in the dilu-
![]()
Figure 1. Relationship between critical N concentration and total biomass with nonlimiting N conditions and total biomass > 1.0 Mg∙DM∙ha−1 according method proposed by Greenwood et al. (1990) for each cultivar during the 2003/4, 2004/5, 2005/6 and 2006/7 growing seasons. IN (▲): Innovator; GR (■): Gem Russet; UR (○): Umatilla Russet; BR (◊): Bannock Russet; MR (+): Markies Russet. The value of coefficients a and b, R2 and p value of allometric function (Nc = a∙W−b) are show in Table 2.
tion N curve depended on the cultivar and was reached before the onset of leaf senescence, when the crops developed their maximum aerial biomass. In Innovator, maximum aerial biomass was reached when the crops produced approximately 15 Mg∙ha−1 of total biomass [13] . The maximum aerial biomass was reached at 19, 20, 24 and 28 Mg∙ha−1 of total biomass production in Gem Russet, Umatilla Russet, Markies Russet and Bannock Russet, respectively (Figure 1). For these last two potato cultivars, the above values were greater than those determined by Greenwood et al. [6] and Duchenne et al. [12] . These results are probably a consequence of the greater yield potential and biomass production of contemporary cultivars [18] [20] [37] .
The values of coefficient a (Table 2) were not significantly different among cultivars and were not significantly different from the coefficients determined by Greenwood et al. [6] , Duchenne et al. [12] and Belanger et al. [14] . Coefficient a represents the plant N concentration when W = 1 Mg∙ha−1 and it depends on the level of steady-state N supply [7] . The values of coefficient b (Table 2) for Innovator and Gem Russet were significantly smaller than those for Umatilla Russet, Bannock Russet and Markies Russet. In addition, the value of coefficient b for Innovator was not significantly different from those reported by Greenwood et al. [6] , Duchenne et al. [12] and Belanger et al. [14] . The value of coefficient b for Gem Russet was significantly different from those reported by Greenwood et al. [6] and Duchenne et al. [12] , and it was not significantly different from those reported by Belanger et al. [14] by the parallelism and the coincidence tests. The values of coefficient b for the other cultivars (Umatilla Russet, Bannock Russet and Markies Russet) were different (p < 0.05) from those determined by these three authors. Bélanger et al. [14] reported different values of coefficients a and similar values of coefficient b for the critical N dilution curve of two potato cultivars (Shepody and Russet Burbank). They argued that the cultivar difference in coefficient a was caused by plant density, as a greater plant density resulted in smaller plants with higher N concentration. Similar b coefficients indicated that both cultivars had similar N dilution. Coefficient b represents the ratio between the relative decline in plant N concentration and the relative crop growth rate [7] . Therefore, the pattern of decrease in N concentration with increasing total biomass was different among cultivars. In summary, the cultivars had different values of coefficient b but showed no differences in coefficient a. The difference in the critical N dilution curve indicates that Innovator requires less N than other cultivars when alL∙have a similar total biomass. This shows that to understand the differences between the dilution coefficients is necessary to study the differences in biomass and N partitioning to tubers of the cultivars.
![]()
Table 2. Coefficients of critical N dilution curve determined for each cultivar and the reference curves. The value in parentheses is the standard error. Data of the 2003/4, 2004/5, 2005/6 and 2006/7 growing seasons were used. The allometric functions are show in Figure 1.
*Significant at the 0.05 probability level; ***Significant at the 0.001 probability level. †coefficients of critical N dilution curve under non-limiting water conditions.
There was a significant interaction between growing seasons (p < 0.005), cultivars and N rates on HI and NHI. Therefore, the variables were assessed within each growing season. The HI and NHI was affected by cultivars and N rates, but without interaction (p > 0.05) in both growing seasons. The HI and NHI were higher in 2005/6 than in 2004/5. Generally, the HI and NHI were significantly greater in Innovator, smaller in Markies Russet and intermediate in the other cultivars (Figure 2). The HI and NHI decreased as the N rate increased, coinciding with the results obtained by Saluzzo et al. [16] and Zebarth et al. [18] . The HI in 2004/5 was higher in N0 (71.4%), lower in N3 (62.5%) and intermediate in treatments N1 and N2 (average 68.7%). Moreover, the HI in 2005/6 was not significantly affected by the N rate (average 77.9%). The NHI was lower in treatments N2 and N3 (average 52.4 and 63.5 for 2004/5 and 2005/6, respectively) than in treatments N0 and N1 (average 58.2 and 69.5 for 2004/5 and 2005/6, respectively). Despite the differences between growing seasons and N rates, the cultivars had different HI and NHI.
Both HI and NHI decreased linearly as the length of the period between planting and maturity of the cultivars increased in both growing seasons (Figure 3). Cultivars with earlier senescence had greater HI and NHI, as previously reported by Kleinkopf et al. [15] and Alva et al. [38] . These results show that Innovator had greater N partitioning to tubers than other cultivars. Also, HI and NHI were significantly correlated with the dilution coefficient (Figure 4). These results demonstrated that the cultivars with higher HI and NHI had greater N dilution. Andriolo et al. [39] reported that potato tubers influenced the N dilution coefficient (b) because of the great contribution of this organ to the total weight of the plant. In the bulking stage, the tubers have a low N concentration, which determines the low content of N in the whole plant. Therefore, after tuber initiation, the dilution coefficient (b) of critical N dilution curve depends on the biomass partitioning to the tubers. Therefore, the differences in HI and NHI among cultivars may explain the differences in the b coefficient of the critical N dilution curve.
3.2. N Nutrition Status
Tuber yield varied between growing seasons and N rates (p < 0.05) in the five cultivars (Table 3). The maximum yields were determined in the treatments with greater N levels and minimum values in the treatment without N. Across all growing seasons, average tuber yield ranged from 43.4 to 63.4 Mg∙ha−1. These values were within the normal range of potato yields for the area where the experiment was conducted [37] . Yield depended on the growth conditions, crop management, and soil N supply, coinciding with the results obtained by Zebarth et al. [2] .
The quadratic model obtained from the relationship between tuber yield and N rate was significant (p < 0.05) (Table 4). Response data for Innovator and Gem Russet in 2003/4 are not included because quadratic coefficient (c) was positive and no statistically significant responses to N rate were observed, respectively. The ONR, ne-
![]()
![]()
Figure 2. Harvest index and nitrogen harvest index for five cultivars in 2004/5 and 2005/6. IN: Innovator; GR: Gem Russet; UR: Umatilla Russet; BR: Bannock Russet; MR: Markies Russet. Vertical lines represent standard error value. The same letter indicates no significant difference among cultivars for each growing season, according to least significant difference (LSD) test (p < 0.05).
![]()
Figure 3. Harvest index and nitrogen harvest index as a function of cultivar maturity (days). Cultivar maturity was assigned a value of 120, 130, 130, 140 and 140 days for Innovator, Gem Russet, Umatilla Russet, Bannock Russet and Markies Russet; respectively. Cultivar maturity = days from planting to crop maturity. Vertical lines represent standard error value.
![]()
Figure 4. Dilution coefficient (b) as a function of harvest index and nitrogen harvest index. r = Correlation coefficient. p = value of probability. Horizontal lines represent standard error value.
![]()
Table 3. Average yield of tuber fresh weight (Mg∙ha−1) for potato cultivars and N rate and mean square and statistical significance for the influence of the growing season and N rate on tuber yield for five cultivars (IN = Innovator; GR = Gem Russet; UR = Umatilla Russet; BR = Bannock Russet; MR = Markies Russet). The same small letter indicates no significant difference among growing season (2003/4, 2004/5, 2005/6 and 2006/7) for each cultivar, according to least significant difference (LSD) test (p < 0.05). The same capital letter indicates no significant difference among treatments (N0, N1, N2 and N3) for each cultivar, according test LSD (p < 0.05).
*Significant at the 0.05 probability level; ***Significant at the 0.001 probability level. nd: not determined.
![]()
Table 4. Quadratic coefficient function for the relationship between yield (tuber fresh weight) and N rate; c: quadratic coefficient; b: linear coefficient; a: intercept, R2: determination coefficient. MY = maximum yield; ONR = optimum nitrogen rate, minimum N rate necessary to achieve maximum yields.
*Significant at the 0.05 probability level; ***Significant at the 0.001 probability level. ns, nonsignificant.
cessary to predict MY, varied between growing seasons for each cultivar, with the values ranging from 150 to 250 kg∙N∙ha−1. This response reflects the fact that the variation in soil N supply can often be as important as crop N demand in determining the optimal fertilizer N rate [2] .
Justes et al. [8] proposed using the NNI to quantify N deficiency or excess. In the present study, for each potato cultivar, values ranged from 0.32 to 1.91 and were similar to those reported by Bélanger et al. [14] in potato (data not shown). In general, during the stages of vegetative growth and tuber initiation (47 and 60 DAP, respectively), the NNI tended to be greater than 1 in all treatments and cultivars, indicating that the crops had no N deficiency. However, at tuber bulking (90 - 100 DAP), the values of NNI were ≤1 in treatments N0 and N1, and ≥1 in treatments N2 and N3. The relationship between RY and NNI varied among cultivars (p < 0.05). According to the parallelism and coincidence tests, the cultivars were separated into two groups: a) Innovator and b) Gem Russet, Umatilla Russet, Bannock Russet and Markies Russet (Figure 5(a)). The RY at 1.0 NNI was 98.0% for Innovator and 99.0% for the other cultivars. The RY decreased with decreasing NNI below 1. Despite its high informative value as diagnostic tool for N crop status, NNI is difficult to use in field conditions and it remains more a research tool than a agricultural management tool. Particularly in this research the NNI allowed to determine differences in plant N status among cultivars. The relationship between RY and dONR also varied with the cultivar (p < 0.05), and agree with the results obtained with NNI (Figure 5(b)). The average RY at zero dONR was 99.6%. These values were near the threshold proposed by Dow and Roberts [40] . Therefore, the model critical N dilution curve and the resulting NNI properly identified the status of N adequately in the potato.
![]() (a) (b)
(a) (b)
Figure 5. Relationship between relative yield (RY) and nitrogen nutrition index (NNI) (a) at tuber bulking (90 - 100 DAP) and N rate differential from the optimum N rate (dONR) (b). IN = Innovator; GR = Gem Russet; UR = Umatilla Russet; BR = Bannock Russet; MR = Markies Russet. Data of the 2003/4, 2004/5, 2005/6 and 2006/7 growing seasons were used. Nitrogen rate differential from the optimum N rate was calculated as the ONR (Table 4) minus the applied N rate within each cultivar and growing season.
At similar N deficiency (NNI < 1; dONR < 0), Innovator had a lower RY than the other cultivars. Innovator showed greater response to similar N rate than the other potato cultivars, but required less N rate to achieve maximum RY. These results show that the potato cultivars had different responses to N fertilization.
4. Conclusion
We conclude that, under the conditions of this study, the processing potato cultivars studied had different critical N dilution curves. The cultivars had different values of coefficient b but showed no differences in coefficient a. Innovator had greater N dilution than the other cultivars. We also determined that the cultivars had different HI and NHI. Innovator had greater HI and NHI than the other cultivars. HI and NHI significantly correlated with coefficient b. Therefore, the differences in HI and NHI among cultivars may explain the differences in coefficient b of the critical N dilution curve. Furthermore, the cultivars had different yield response as the NNI decreased. At similar N deficiency, Innovator had lower RY and greater response to N rate than the other potato cultivars. Therefore, the critical N dilution curve can be used to define the N requirements during growing season and the NNI identify situations of deficient and non-deficient N nutrition adequately in potato cultivars. Critical N dilution curve and NNI are research tools and these allow determining differences in plant N status among potato cultivars.
Acknowledgements
This work is part of a thesis submitted by Claudia Giletto in partial fulfillment for the requirements for the PhD degree of the Universidad Nacional de Mar del Plata, Buenos Aires, Argentina. This study was supported by the INTA project PNHFA-110607and FCA-UNMP AGR 474/15.
Abbreviations
BR, Bannock Russet;
DAP, Days after Planting;
dONR, Nitrogen Rate Differential from the Optimum Nitrogen Rate;
GR, Gem Russet;
HI, Harvest Index;
IN, Innovator;
MR, Markies Russet;
N, Nitrogen;
NHI, Nitrogen Harvest Index;
Nc, Critical Nitrogen Concentration;
NNI, Nitrogen Nutrition Index;
ONR, Optimum Nitrogen Rate;
RY, Relative Yield;
UR, Umatilla Russet.
NOTES
![]()
*Corresponding author.