Nonlinear Evolution Equations and Its Application to a Tumour Invasion Model ()
1. Introduction
In this paper we consider the initial Neumann-boundary value problem of nonlinear evolution equations with logistic term, arising from tumour invasion models with proliferation and re-establishment: (NE)
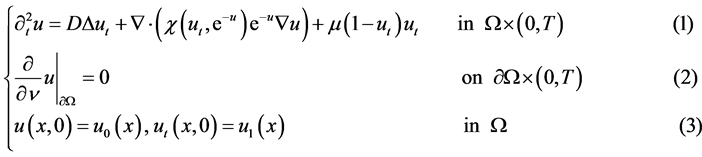
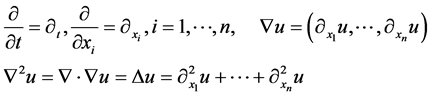
where  for
for , D and μ are positive constants, Ω is a bounded domain in
, D and μ are positive constants, Ω is a bounded domain in  and
and 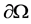 is a smooth boundary of Ω and
is a smooth boundary of Ω and 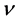 is the outer unit normal vector.
is the outer unit normal vector.
Let us introduce function spaces used in below. First, 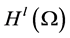 denotes the usual Sobolev space
denotes the usual Sobolev space 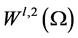 of order l on Ω. For functions
of order l on Ω. For functions 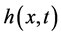 and
and 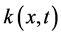 defined in
defined in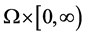 , we denote
, we denote
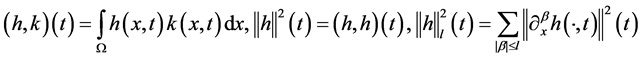
where 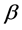 is a multi-index for
is a multi-index for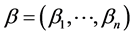 .
.
The eigenvalues of 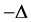 with the homogeneous Neumann boundary conditions are denoted by
with the homogeneous Neumann boundary conditions are denoted by 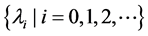 satisfying
satisfying 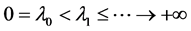 and
and ![]() indicates the
indicates the ![]() normalized eigenfunction corresponding to
normalized eigenfunction corresponding to![]() . For a non-negative integer l, we set
. For a non-negative integer l, we set ![]() as a closure of
as a closure of ![]() in the function space
in the function space![]() . It is noticed that we have
. It is noticed that we have ![]() for
for![]() , which enables us to use Poincare’s Inequality.
, which enables us to use Poincare’s Inequality.
Putting ![]() in (1) it follows from (1) that
in (1) it follows from (1) that
![]()
where ![]() and
and ![]() are positive parameters. (NE) is rewritten by the following problem.
are positive parameters. (NE) is rewritten by the following problem.
![]()
where ![]() We will show the global existence in time of solutions of (RP), which gives our desired result of (NE).
We will show the global existence in time of solutions of (RP), which gives our desired result of (NE).
Applying the above result to the following mathematical model of tumour invasion proposed by Chaplain and Lolas [1] , we have a rigorous mathematical understanding to tumour invasion for the key variables![]() ,
, ![]() and
and![]() .
.
![]()
where ![]() is the density of tumour cells,
is the density of tumour cells, ![]() is matrix degradation enzymes (MDEs) concentration and
is matrix degradation enzymes (MDEs) concentration and ![]() is extracellular matrix (ECM) density in
is extracellular matrix (ECM) density in ![]() and
and![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() and
and ![]() are positive constants. In the right hand side of (4) the second, third and fourth terms mean chemotaxis, haptotaxis and proliferation of tumour cell respectively. Also the second term of the right hand side of (5) describes the re-establishment of ECM. We consider an initial boundary value problem for (C-L) satisfying
are positive constants. In the right hand side of (4) the second, third and fourth terms mean chemotaxis, haptotaxis and proliferation of tumour cell respectively. Also the second term of the right hand side of (5) describes the re-establishment of ECM. We consider an initial boundary value problem for (C-L) satisfying
![]()
![]() ,
,
where ![]() are initial data of
are initial data of![]() .
.
Chaplain and Anderson [2] , corresponding to the case of ![]() in (C-L), proposed a mathematical model on generic solid tumour growth at the avascular stage. While most tumours are asymptomatic at this stage, it is still possible for cells to escape and migrate to the lymph nodes and for more aggressive tumours to invade.
in (C-L), proposed a mathematical model on generic solid tumour growth at the avascular stage. While most tumours are asymptomatic at this stage, it is still possible for cells to escape and migrate to the lymph nodes and for more aggressive tumours to invade.
In our previous papers [3] [4] , we consider only the case of ![]() and
and ![]() for our convenience. In this paper we may consider the case where
for our convenience. In this paper we may consider the case where ![]() is sufficiently small positive constant. Actually compared with the effect of haptotaxis, the one of chemotaxis is very small. Furthermore we study the case of
is sufficiently small positive constant. Actually compared with the effect of haptotaxis, the one of chemotaxis is very small. Furthermore we study the case of![]() , which describes tumour invasion phenomena with tumour cell proliferation and re-establishment of ECM respectively.
, which describes tumour invasion phenomena with tumour cell proliferation and re-establishment of ECM respectively.
In the final section by computer simulations of our model (by Mathematica 8) we can easily observe time-dependent interaction and the relationship between the above components in complicated procedure of tumour invasion and a comparison to our rigorous mathematical result. Comparing our mathematical result with computer simulations we will gain a better understanding of the mechanism of tumour invasion.
2. Existence Theorem of (NE)
By deriving the energy estimate of (RP) (see [3] - [9] ) and considering the iteration scheme we obtain existence of solutions to (RP) by the standard argument to show the convergence of solutions of the iteration scheme.
In the same way as used in [3] [4] [5] [6] [7] we have the following estimates of (RP). We begin with ![]() in order to obtain a basic estimate of (RP). Then we have for
in order to obtain a basic estimate of (RP). Then we have for![]() ,
,
![]()
by the integration by parts
![]() (7)
(7)
It is noticed that the following estimate is obtained in [4] [5] [6] [7] for![]() ,
,
![]() (8)
(8)
Then for the nonlinear term we have by using (8)
![]() (9)
(9)
where we used Dionne [10] for the estimate of nonlinear terms and ![]() is sufficiently small positive constant. Therefore we have by integrating the both sides of (7) over
is sufficiently small positive constant. Therefore we have by integrating the both sides of (7) over ![]() and using (9)
and using (9)
![]() (10)
(10)
where we denote ![]()
Since the last term of the right hand side of (10) is negligible for sufficiently small![]() , we have by integration by parts with respect to t
, we have by integration by parts with respect to t
![]() (11)
(11)
Taking ![]() sufficiently large for the second term of the right hand side of (11) is negligible. Since
sufficiently large for the second term of the right hand side of (11) is negligible. Since ![]() if
if![]() , the third term in the left hand side of (11) is positive. Then we obtain a basic energy estimate
, the third term in the left hand side of (11) is positive. Then we obtain a basic energy estimate
![]() (12)
(12)
Replacing![]() ,
, ![]() instead of
instead of ![]() in the above procedure, we obtain the following estimate of higher order.
in the above procedure, we obtain the following estimate of higher order.
Lemma 1 (Energy estimate of (RP)) Assume that ![]() for
for ![]() satisfies appropriate smooth regularity condition. Supposed that
satisfies appropriate smooth regularity condition. Supposed that ![]() we have the energy estimate of (RP) for
we have the energy estimate of (RP) for ![]()
![]() (13)
(13)
where we denote for any non-negative integer![]() ,
, ![]()
We consider the iteration scheme of (RP):
![]()
where
![]()
(13) guarantees the uniformly bounded estimate of ![]() for
for![]() . We determine
. We determine ![]() by the solution of the following ordinary equations with initial data.
by the solution of the following ordinary equations with initial data.
![]()
The local existence in time of ![]() is shown by the theory of ordinary differential equations. Therefore, by the energy estimates, the global existence in time of the solution
is shown by the theory of ordinary differential equations. Therefore, by the energy estimates, the global existence in time of the solution ![]() satisfying the regularity required for Lemma 1 and justification of the limiting process are assured by the standard method. The energy estimate enables us to get the solution of (RP) in
satisfying the regularity required for Lemma 1 and justification of the limiting process are assured by the standard method. The energy estimate enables us to get the solution of (RP) in
![]()
by considering ![]() and standard argument of convergence for
and standard argument of convergence for ![]() (see [3] - [9] ).
(see [3] - [9] ).
Then we obtain the following result of (NE) by using the above result of (RP).
Theorem 2 Assume that ![]() for
for ![]() satisfies appropriate smooth regularity condition, initial data
satisfies appropriate smooth regularity condition, initial data ![]() are sufficiently smooth for
are sufficiently smooth for![]() ,
, ![]() and that
and that ![]() is small enough. For sufficiently large
is small enough. For sufficiently large![]() , there is a solution for
, there is a solution for ![]()
![]()
to (NE) such that it satisfies the following asymptotic behavior
![]()
3. Application to a Tumour Invasion Model
Their mathematical approach to above models mainly depends on numerical analysis. In this paper first we show the rigorous mathematical result of (C-L) and then computer simulations, of which the validity is guaranteed by our mathematical result.
On the other hand, there are many mathematical models which can be found in the literature describing tumour angiogenesis. In [20] Levine and Sleeman applied the mathematical model of Othmer and Stevens [21] for the understanding of tumour angiogenesis, which arises in the theory of reinforced random walk. Anderson and Chaplain [12] proposed a model of tumour angiogenesis taking account of endothelial tip-cell migration. The model describes cell migration governed by three factors: diffusion, chemotaxis and haptotaxis.
Rigorous Mathematical approaches to tumour growth models have been known (see [3] - [9] [20] [21] [22] [23] [24] ). Levine and Sleeman [20] and Yang, Chen and Liu [24] studied the global existence in time of solutions and blow up ones to a simplified Othmer and Stevens model. Kubo et al. [3] - [9] show the time global solution and asymptotic behavior of the solution to the mathematical models proposed by [2] [12] [20] [21] [23] .
3.1. Reduced Problem
Following to Levine and Sleeman [20] we reduce our problem to a simpler system (see [3] - [9] [20] ). It is easily seen in (5) that ![]() is written by
is written by
![]() (14)
(14)
Integrating (14) over ![]() for
for ![]()
![]()
Put ![]() and
and ![]() for a constant
for a constant![]() . Then we have
. Then we have
![]()
denoting ![]() and
and ![]()
![]() (15)
(15)
Substituting ![]() by the right hand side of (14), from (4) and (6) it follows that for
by the right hand side of (14), from (4) and (6) it follows that for ![]()
![]() (16)
(16)
and
![]() (17)
(17)
The nonlinear evolution Equation (1) involves (16) and so we can apply Theorem 2 to (16).
3.2. Existence Theorem of (C-L) with ![]()
The Equations (16) and (17) are essentially regarded as the same type of equation as (1). Hence the energy estimates of ![]() and
and ![]() are obtained and combining these estimates we obtain the desired estimate (cf. [3] [4] [5] [6] [7] ).
are obtained and combining these estimates we obtain the desired estimate (cf. [3] [4] [5] [6] [7] ).
Lemma 3 (Energy estimate of (C-L)) We obtain the energy inequality of the reduced problem (16) and (17) with zero-Neumann boundary condition for ![]() and sufficiently large
and sufficiently large ![]()
![]() (18)
(18)
where ![]() as
as![]() .
.
Then applying the same argument as used for Theorem 2 to the above mathematical model, we obtain global existence in time and asymptotic behaviour of the solutions to our mathematical model.
Our main result for ![]() is as follows.
is as follows.
Theorem 4 For smooth initial data ![]() assume that
assume that ![]() - norms of
- norms of![]() ,
, ![]() and
and ![]() are sufficiently small, then there are classical solutions of (C-L) for
are sufficiently small, then there are classical solutions of (C-L) for![]() :
: ![]() such that they satisfy the following asymptotic behavior for
such that they satisfy the following asymptotic behavior for ![]()
![]()
Also we can deal with the case of ![]() as follows.
as follows.
3.3. Existence Theorem of (C-L) with ![]()
In ![]() for
for![]() , especially we may consider
, especially we may consider ![]() -inner product composed of the chemotaxis term and
-inner product composed of the chemotaxis term and ![]()
![]()
in the same way as in Section 2
![]()
where we can take ![]() small enough for sufficiently small
small enough for sufficiently small![]() . It is noticed that compared with the effect of haptotaxis, the one of chemotaxis may be quite small. Hence since we can have the same estimate as Lemma 3 for sufficiently small
. It is noticed that compared with the effect of haptotaxis, the one of chemotaxis may be quite small. Hence since we can have the same estimate as Lemma 3 for sufficiently small![]() , the same result as in Theorem 4 holds for this case too.
, the same result as in Theorem 4 holds for this case too.
Theorem 5 Under the same assumption as in Theorem 4 we further assume that ![]() is sufficiently small, then we obtain the same result as stated in Theorem 4.
is sufficiently small, then we obtain the same result as stated in Theorem 4.
4. Computer Simulations
In Kolev and Zubik-Kowal [16] the same type model of (C-L) for ![]() was considered and they obtained numerical solutions. In this paper our advantage is to show global existence in time of rigorous mathematical smooth solutions of the model and based on it carry out computer simulations.
was considered and they obtained numerical solutions. In this paper our advantage is to show global existence in time of rigorous mathematical smooth solutions of the model and based on it carry out computer simulations.
In the following Figures 1-7 illustrated below we show the graphs along the time by the computer simulations of the model at ![]() and 1.35 by Mathematica 8. We use the parameter values of
and 1.35 by Mathematica 8. We use the parameter values of![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() and
and ![]() specified below.
specified below.
We will observe the time dependent relationship and interaction between tumour cells (Red line), the surrounding tissue (Black line) and degradation enzymes (Green line). In the graphs below a coordinate axis of the horizontal direction indicates the spatial position and vertical direction indicates the density or concentration of each component of the model.
![]()
Figure 1. Interactions between the tumour and the surrounding tissue without proliferation of tumour cell, migration, and ECM re-establishment: The parameter values dn = 0.001, dm = 0.001, γ = 0.02, η = 10, μ1 = 0, μ2 = 0, α = 0 and β = 0.1, xn = 0. We can observe that MDEs degradates the surrounding tissue, and makes space into which tumour cells move. Further tumour cells form a small peak and keep going forward inside ECM, preserving the shape.
![]()
Figure 2. Tumour cell proliferation, migration, and interactions between the tumour and the surrounding tissue without ECM re-establishment: The parameter values dn = 0.001, dm = 0.001, γ = 0.02, η = 10, μ1 = 5, μ2 = 0, α = 0.1 and β = 0.1, xn = 0. It is seen that tumour cell density becomes much higher than in Figure 1 inside ECM and maintains the upper bound of the logistic curve constantly.
![]()
Figure 3. Tumour cell proliferation, migration, without ECM re-establishment, and interactions between the tumour and the surrounding tissue: The parameter values dn = 0.001, dm = 0.001, γ = 0.02, η = 10, μ1 = 10, μ2 = 0, α = 0.1 and β = 0.1, xn = 0. Increasing μ1 more, it is observed that tumour cell density inside ECM become higher than in Figure 2.
![]()
Figure 4. Tumour cell proliferation, migration, ECM re-establishment, and interactions between the tumour and the surrounding tissue: The parameter values dn = 0.001, dm = 0.001, γ = 0.02, η = 10, μ1 = 5, μ2 = 0.001, α = 0.1 and β = 0.1, xn = 0. We take μ2 = 0.001 only and other parameters are same as in Figure 2. Then the tumour cell density is almost same as in Figure 2.
![]()
Figure 5. Tumour cell proliferation, migration, ECM re-establishment, and interactions between the tumour and the surrounding tissue: The parameter values dn = 0.001, dm = 0.001, γ = 0.02, η = 10, μ1 = 5, μ2 = 1, α = 0.1 and β = 0.1, xn = 0. When taking μ2 = 0.001, compared with Figure 2 and Figure 4, it is clear that the invasive area of tumour cells inside ECM is much more limited.
![]()
Figure 6. Tumour cell proliferation, migration, ECM re-establishment, and interactions between the tumour and the surrounding tissue: The parameter values dn = 0.001, dm = 0.001, γ = 0.02, η = 10, μ1 = 5, μ2 = 5, α = 0.1 and β = 0.1, xn = 0. Compared with Figure 4 and Figure 5, the reachable range of tumour cells is observed to be more suppressed by taking μ2 = 5.
![]()
Figure 7. Tumour cell proliferation, chemotaxis, migration, ECM re-establishment, and interactions between the tumour and the surrounding tissue: The parameter values dn = 0.001, dm = 0.001, γ = 0.02, η = 10, μ1 = 5, μ2 = 0, α = 0.1 and β = 0.1, xn = 0.01. By the effect of chemotaxis (xn = 0.01), tumour cells are attracted by MDEs, the density is beyond 1 at t = 0.25 ~ 0.5, eventually it converges to 1 and after that keeps it constantly.
5. Conclusions
In order to obtain the global existence in time and asymptotic profile of solutions of a mathematical model of tumour invasion proposed by Chaplain and Lolas, we investigate nonlinear evolution equations with logistic term related to our mathematical models as an initial Neumann-boundary value problem. We could show the global existence in time of rigorous mathematical solutions to the initial boundary value problem for the model in arbitrary space dimension by using the energy inequalities. Applying the result to our model we show global existence in time of mathematical solutions of the model.
By Figures 1-7, it is recognized that our rigorous mathematical result of the existence and asymptotic behaviour of smooth solutions verifies our computer simulations and confirms the pattern form of each component of the model in the graphs respectively. Then we can gain the understanding of the process of tumour invasion more in details.
Acknowledgements
This work was supported in part by the Grants-in-Aid for Scientific Research (C) 19540200, 22540208, 25400148 and 16K05214 from Japan Society for the Promotion of Science.