Genetic Worth Effect Models for Boreal Conifers and Their Utility When Integrated into Density Management Decision-Support Systems ()
1. Introduction
Black spruce (Picea mariana (Mill.) BSP) and jack pine (Pinus banksiana Lamb.) are among the most preferred reforestation species within the central portion of the Canadian Boreal Forest Region (Rowe, 1972) given their current ecological suitability and commercial importance. In the Province of Ontario, black spruce and jack pine constitute approximately 35% and 12% of the total growing stock, respectively (Watkins, 2011) . The nominal silvicultural prescription employed in reforesting upland black spruce and jack pine site types under an intensive or elite management intensity following a stand replacing disturbance, such as clear cut harvesting, includes: 1) mechanical site preparation followed by planting of genetically enhanced stock; 2) controlling inter specific competition via vegetative management treatments during the pre-crown closure stage of development; and 3) density control treatments consisting of commercial thinning treatments in order to reduce mortality losses during the post-crown closure stage of development. Specific treatment decisions regarding initial spacing and thinning treatments for a given site, management intensity and volumetric objective, are commonly determined using stand density management decision-support models (McKinnon et al., 2006) .
Recently, however, the complexity of designing optimal crop plans has become more challenging as management strategies attempt to move away from an objective strictly concerned with the maximization of volumetric yield to one that also addresses end-product quality and value maximization goals (Emmett, 2006) . Fortunately, the complexity of such crop planning has been greatly reduced with the advent of structural stand density management models (SSDMMs; Newton, 2009, 2012a ). Briefly, SSDMMs are distance-independent diameter-distribution yield models that are derived from the modeling platform used to develop stand density management diagrams (SDMDs). Since their inception in Japan in the 1960s (e.g., Ando, 1962, 1968 ), SDMDs have evolved over time in terms of their complexity and utility: progressing from static (Ando, 1962; Drew & Flewelling, 1979; Archibald & Bowling, 1995) to dynamic (Newton & Weetman (1993, 1994); Stankova & Shibuya (2006)) to structural (Newton et al. (2004, 2005)) models. These ecological-based crop planning tools have been developed for numerous forest tree species throughout the boreal, temperate and tropical forest regions (Drew & Flewelling, 1977; Jack & Long, 1996; Newton, 1997) . One area where SDMDs have found the greatest affinity is in Japan where regional-specific variants have been developed for most of their commercial important and intensely managed coniferous stand types (e.g., Forestry Agency of Japan, 1981a, 1981b; Fujimori, 2001 ).
Commonly, SSDMMs attempt to account for various silvicultural interventions such as thinning and genetic worth effects through adjustments to the underlying driving functions; principally, through the species and site specific mean dominant height-age functions (e.g., Newton, 2012a ). These modeling adjustments account for the accelerated rate of stand dynamics that frequently arise from the enhanced growth performance of individual trees following treatment. Conceptually, these approaches belong to the treatment modifier equation class of methodologies use for modeling growth responses to silvicultural treatments (Weiskittel et al., 2011) . Currently, however, the genetic worth effect models within the SSDMMs do not account for the expected temporal decay of genetic gains over time (sensu Lambeth, 1980 ). Consequently, in order to improve the predictability of SSDMMs and advance genetic worth effect modelling within the context of SDMD-based decision-support systems, the objectives of this study were to introduce a modeling solution for addressing this shortcoming and subsequently integrated it into the SSDMM architecture. The proposed solution follows established practice and is consistent with previous approaches use to model genetic worth effects (e.g. Xie & Yanchuk, 2003 ). Operationally-relevant crop planning simulations are used to demonstrate the utility of the revised SSDMMs.
2. Material and Methods
2.1. Approach
Conceptually, genetic worth effect response models attempt to quantitatively account for the permanent acceleration in stand development which arises from the use of genetically enhanced stock. Consequently, the approach utilized in this study consisted of modelling genetic worth effects through an empirical-based adjustment to the species and site specific mean dominant height-age function, which is the principal driver of temporal change within not only SSDMMs but also previous generations of SDMD-based models. Two response model variants were considered: one in which the increase rate of stand development initiates at the specified genetic- based selection age and then dissipates over time in a linear fashion according to an empirical juvenile age-mature age correlation model (Type 1 response) ; and the other in which the increase initiates at the time of plantation establishment and continues until the specified selection age, and thereafter dissipates according to an empirical juvenile age-mature age correlation model (Type 2 response). This approach assumes that for a given species, generation, region or seed lot, the genetic worth effect is known in terms of the expected height growth increase and the corresponding selection age. This information is commonly available from the results of previous tree improvement experimentation including interprovincial trials and meta-analytical summaries (e.g., Newton, 2003 ).
2.2. Model Formulation
The Type 1 response model expresses the genetic worth effect in terms of the percentage increase in dominant
height  at the specified selection age (Equation (1)) and then embeds this increase within the
at the specified selection age (Equation (1)) and then embeds this increase within the
site-specific height-age model.
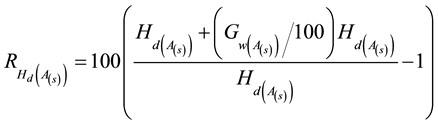 (1)
(1)
where  and
and 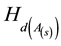 are the specified genetic worth effect (%) at the specified selection age (A (s) (yr))
are the specified genetic worth effect (%) at the specified selection age (A (s) (yr))
and mean dominant height (m) at A (s) as predicted by site-index-based height-age models (e.g., models developed for upland black spruce by Carmean et al. (2006) and jack pine by Carmean et al. (2001) ). This increase is then used to adjust the height-age estimates initiating at A(s) and proceeding to the specified rotation age, according to the following genetic-based juvenile age-mature age correlation model which was previously developed for Pincaceae ( Lambeth, 1980 ; Equation (2)).
 (2)
(2)
where  is the relative percentage increase in height growth at the ith age (A(i)).
is the relative percentage increase in height growth at the ith age (A(i)).
The Type 2 response model first predicts a backwards predicted gain from the time of establishment to
A(s) according to the specified  and the age-age correlation model ( Lambeth, 1980 ; Equation
and the age-age correlation model ( Lambeth, 1980 ; Equation
(3)).
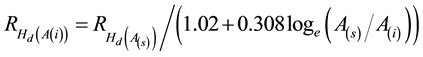 (3)
(3)
Subsequently from the selection age, the increase is thereafter estimated using the juvenile age-mature age correlation relationship (Equation (2)). The Type 1 and 2 response models predict equivalent height growth percentage gains from the specified selection age until rotation. However, they predict different stand development rates during the early phase of plantation development, specifically, from establishment until the plantation reaches the specified selection age.
2.3. Integration into SSDMMs
For a given stand-type, site quality and crop plan which includes specific information regarding the initial density, genetic worth effects, thinning treatments, merchantable specifications, operational adjustment factors, degrade factors, cost profiles, and rotation length, the enhanced modular-based SSDMMs employ a hierarchical computation sequence to estimate volumetric yields, diameter distributions, tree heights, log assortments, components-specific biomass and carbon outcomes, sawmill-specific products and associated values, and fibre quality attributes (e.g., see Newton, 2012a ). Within the SSDMM structure, the new response models for genetic worth effects were integrated within Module A given that this module includes all the core relationships that predict the temporal patterns of stand development (e.g., see Newton, 2012a ). The revised computational sequence is described in the following schematic-based script.
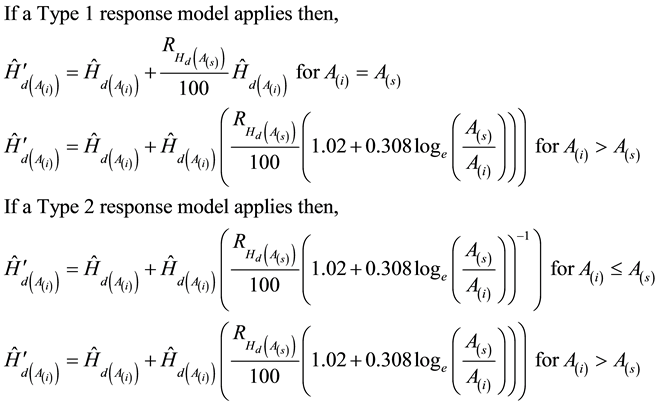
where  is redefined as
is redefined as 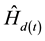 after each age increment.
after each age increment.
3. Results
The genetic worth effect models for black spruce and jack pine plantations are exemplified in Figure 1 using
![]()
Figure 1. Contrasted site-specific dominant height-age curves for black spruce (left hand side) and jack pine (right hand side) plantations established on good site qualities (site index of 18 m) with the embedded Type 1 and Type 2 genetic worth effect response model which employed a 10% worth effect at a selection age of 15 yr for the black spruce plantations and a 7% worth effect at a selection age of 20 yr for the jack pine plantations (filled circle), versus corresponding plantations established with non-improved stock (open circle).
meta-based estimates of genetic gain for a given site quality. Specifically, the modified site-specific mean dominant height-age relationships arising for both response types are contrasted with that projected for unimproved stock by species. For an expected genetic gain value of 10% at a selection age of 15 yr for black spruce and 7% at 20 yr for jack pine (Newton, 2003) , plantations established on good site qualities (site index of 18 m) and managed to a rotation age of 60 yr, the Type 1 response model would predict a dominant height increase of 10% (from 5.1 to 5.6 m) at age 15 yr for black spruce, and 7% (from 8.9 to 9.6 m) at age 20 yr for jack pine, according to Equation (1). These relative increases would then decline in a linear pattern according to the juvenile age-mature age correlation relationship (Equation (2)). Similarly, the predicted genetic worth effect according to the Type 2 response model would result in increased height values from ages 1 to 15 yr for black spruce, and from ages 1 to 20 yr for jack pine (Equation (1)), maximizing at age 15 yr for black spruce and 20 yr for jack pine, according to the specified genetic worth effect and selection age provided. Thereafter the increases would decline according to the juvenile age-mature age correlation relationship (Equation (3)). As expected, the Type 1 and 2 response models predicted equivalent rotational heights (20.2 m and 20.1 m for black spruce and jack pine, respectively), rotational heights were greater for the plantations established using improved stock compared to the plantations composed of unimproved stock (e.g., 6% and 5% for black spruce and jack pine, respectively), and relative height gains dissipated over time from 10% at age 15 yr to 6% at age 60 yr for black spruce, and from 7% at age 20 yr to 5% at age 60 yr for jack pine.
Furthermore, extending these examples within the context of the revised SSDMMs enables one to assess of the differences between the response models in terms of rotational yields and productivity indices. Briefly, the hierarchical-based SSDMMs consisted of 6 sequentially-linked estimation modules (Newton, 2009, 2012a) : Module A-Dynamic SDMD; Module B-Diameter and Height Recovery; Module C-Taper Analysis and Log Estimation; Module D-Biomass and Carbon Estimation; Module E-Product and Value Estimation; and Module F- Fibre Attribute Estimation. Module A is a mathematical representation of a dynamic stand density management diagram and consists of a broad array of static and dynamic yield―density relationships. Module B consists of 1) Weibull-based parameter prediction equation systems for diameter distribution recovery, and 2) composite height-diameter prediction equations for diameter-class-specific height estimation. Module C employs a dimensional compatible taper equation to predict log products (number of pulp and saw logs) and stem volumes of individual trees. Module D includes allometric-based composite biomass equations for each above-ground component (bark, stem, branch and foliage) from which diameter-class and stand-level biomass and associated carbon-based equivalents are predicted. Module E utilizes sawmill-specific (stud and random length mill) product and value equations to predict diameter-class and stand-level chip and lumber volumes and associated monetary values. Module F contains composite equations for estimating wood density and mean maximum branch diameter. A schematic illustration of the structure of the SSDMM including the interrelationships and sequential flow of computations among the individual modules is provided in Newton (2012a) . The algorithmic analogue of these SSDMMs was developed so that a set of crop plans could be simultaneously contrasted and assessed employing a comprehensive set of yield outcome metrics and performance measures. These measures included indices for quantifying overall productivity, log quality and product distributions, biomass production and carbon yields, quantity and value of recoverable end-products, economic efficiency, duration of optimal site occupancy, structural stability, quality of fibre attributes, and operability status.
The input settings for the simulations used to demonstrate the utility of the genetic worth effect models were as follows. For each species, 3 plantations established at initial densities of 2500 stems/ha on good quality sites (site index of 18 m at 50 yr breast-height age) were grown to a rotation age of 60 yr. The control stand consisting of unimproved stock (no genetic worth effects included) is represented by Regime 1, the plantation employing the Type 1 genetic worth effects response model which accounts for the increased rate of development during the post selection age period is represented by Regime 2, and plantation employing the Type 2 genetic worth effects response model which accounts for increased rate of development over the entire rotation is represented by Regime 3. For each of the 3 regimes, a fixed cost of $0.3K/ha was used to cover regeneration assessment and site preparation costs at the time of plantation establishment. Planting costs were set at $0.8 per planted seedling. Density-independent mortality throughout the rotation as quantified by the operational adjustment factor was set to 0.01%/yr. Genetic worth and selection age were set at nominal values of 10% and 15 yr for black spruce and 7% at 20 yr for jack pine (Newton, 2003) . Rotational variable costs associated with stumpage and renewal fees, harvesting, transportation and manufacturing were set at a constant value ($75/m3). The time of simulation, rate of inflation and discount rate were set to 2014, 2% and 4%, respectively, and a 10% degrade factor for adjusting product values were used. Reflecting a sawlog management objective in which there is a bias towards the production of larger but few trees at rotation, the operability targets were set at a piece size and merchantable volume yield threshold of 10 stems/m3 and 200 m3/ha, respectively.
The temporal mean volume-density trajectories for these regimes within the context of the traditional SDMD mean volume-density graphic are illustrated in Figure 2(a) for black spruce and Figure 2(b) for jack pine. The
plantations composed of genetically enhanced stock (Regimes 2 and 3) attained higher levels of site occupancy irrespective of species or response model type, as evident by their greater mean size-density condition at rotation. Table 1 presents the resultant rotational yield estimates and Table 2 provides the rotational productivity and performance metrics by species for plantations composed of unimproved stock versus those composed of genetically improved stock. Contrasting the yield estimates for plantations established with unimproved stock versus those established with genetically enhanced stock, indicated that the plantations established using improved stock had 1) greater rotational mean tree sizes (height (6% for black spruce and 5% for jack pine), diameter (4% and 5%) and volume (20% and 17%)), 2) attained higher levels of stocking and site occupancy (basal area (6% and 6%) and relative density index (4% and 8%)), 3) produced greater total volumes (11% and 4%) and merchantable volume (12% and 11%), fewer number of pulplogs (−3% and −5%) but a greater number of sawlogs (13% and 34%), (4) greater recoverable products (chip (6% and 9%) and lumber (21% and 16%)) and higher economic worth (26% and 30%). In terms of performance measures, differences among the regimes indicated that the plantations composed of improved stock were 1) more productive in terms of volume (12%
![]()
Table 1. Rotational yield estimates for black spruce and jack pine plantations established using unimproved (Regime 1) and genetically enhanced (Regimes 2 and 3) stock.
aMerchantable volume specifications: 2.59 m and 5.03 m pulp and sawlog length, respectively, 4 cm merchantable top, 10 cm and 14 cm minimum log diameter for pulp and saw logs, respectively. S and R refer to stud and randomized length mill configurations, respectively. See Newton (2012a) for a complete computational summary of these metrics. bTreatment specifics given in the text.
![]() (a)
(a)![]() (b)
(b)
Figure 2. (a) SDMD graphic for black spruce plantations: (i) isolines for mean dominant height (Hd; solid horizontal lines: 4 m (bottom) - 20 m (top) by 2 m intervals), quadratic mean diameter (Dq; long-dashed horizontal lines; 4 cm (bottom) - 26 cm (top) by 2 cm intervals), mean live crown ratio (Lr; short-dashed diagonal lines; 35% (bottom), 40%, 50%, …, 80% (top)), and relative density index (Pr; dotted diagonal lines; 0.1 - 1.0 by 0.1 intervals); (ii) crown closure line (left-side solid diagonal boundary line) and self-thinning rule at a Pr = 1.0 (right-side solid diagonal boundary line); (iii) lower and upper Pr multi-dashed diagonal lines delineating the optimal density management window (Dm; 0.32 ≤ Pr ≤ 0.45); and (iv) expected 60 year size-density trajectories with 1 year intervals denoted by a vertical line for each of the 3 user-specified crop plans for plantations situated on good quality sites (SI = 18). Specifically: Regime 1―initial planting density of 2500 stems/ha with no thinning employing the null genetic response model; Regime 2―establishment density of 2500 stems/ha with no thinning employing the Type 1 genetic response model; and Regime 3―establishment density of 2500 stems/ha with no thinning employing the Type 2 genetic response model; (b) SDMD graphic for jack pine plantations: (i) isolines for mean dominant height (Hd; solid horizontal lines: 4 m (bottom) - 22 m (top) by 2 m intervals), quadratic mean diameter (Dq; long-dashed horizontal lines; 4 cm (bottom) - 26 cm (top) by 2 cm intervals), mean live crown ratio (Lr; short-dashed diagonal lines; 35% (bottom), 40%, 50%, …, 80% (top)), and relative density index (Pr; dotted diagonal lines; 0.1 - 1.0 by 0.1 intervals); (ii) crown closure line (left-side solid diagonal boundary line) and self-thinning rule at a Pr = 1.0 (right-side solid diagonal boundary line); (iii) lower and upper Pr multi-dashed diagonal lines delineating the optimal density management window (Dm; 0.32 ≤ Pr ≤ 0.45); and (iv) expected 60 year size-density trajectories with 1 year intervals denoted by a vertical line for each of the 3 user-specified crop plans for plantations situated on good quality sites (SI = 18). Regimes specifics follow those given in Figure 2(a).
![]()
Table 2. Stand-level performance indices for black spruce and jack pine plantations established using unimproved (Regime 1) and genetically enhanced (Regimes 2 and 3) stock.
aMAVI, MABI and MACI denote mean annual merchantable volume, biomass and carbon increment, respectively. S and R refer to stud and randomized length mill configurations, respectively. See Newton (2012a) for a complete computational summary of these metrics; bAs described in the text.
and 11%), biomass (14% and 9%), and carbon sequestration (14% and 6%) production, 2) produced a greater proportion of usable end-products (saw logs (6% and 29%) and lumber volume (5% and 2%)), and 3) attained operability status at an earlier age (−8% and −9%). Stand stability (height/diameter ratio), wood density and maximum branch size differences were inconsequential among the regimes for both species.
Comparing the Type 1 and 2 response models revealed that the rotational outcomes were approximately equivalent within each species. This is in accord with expectation given that both response models constrain mean dominant height to be equivalent at the specified selection age and thereafter proceed along approximately identical height-age pathways until rotation. The only real difference relates to the early phase of plantation development: i.e., from establishment and until the specified selection age is reached. During this period, the Type 2 response model predicts a greater rate of development than does the Type 1 response model given that the genetic worth effect initiates at the time of plantation establishment rather than at the specified selection age. Hence, the Type 2 response model attained greater mean dominant heights for each year before the selection age was reached, resulting in a slight increase in the rate of stand development. As a consequence, plantations growing according to the Type 2 response model would attain crown closure status slightly earlier than plantations growing according to the Type 1 response model.
4. Discussion
Conceptually, the modelling approach presented in this study is similar to the treatment modifier equation class of methodologies described by Weiskittel et al. (2011) . Specifically, by revising existing untreated prediction models by adding rate parameter modifiers in order to reflect treatment-induced changes to stand development rates. Various approaches used in modelling the growth and yield consequences arising from silvicultural treatments and interventions have been proposed. These include parameterization of treatment-specific response models, multivariate functions with treatment variables explicitly included in the model specification, and the treatment modifier method as used in this study. Among these approaches, the treatment modifier approach has been the most frequently applied. This is principally due to the fact that existing equations which drive the overall dynamics of growth and yield simulators can be readily modified without the need of new data acquisition, model specification and parameterization initiatives (Weiskittel et al., 2011) .
Once the treatment modifier equation method has been selected and the expected effects of the treatments are known in terms of their magnitude, duration and temporal initiation, existing models can be modified accordingly. For genetic worth effects, both the Type 1 and 2 response models were included given that genetic gains are long-term in their duration and affect stand dynamics over most or all of the rotation. The presented response models differentiate themselves in this regard: Type 1 is applicable from the specified selection age until rotation age whereas the Type 2 is applicable from the time of establishment until rotation age. However, their effects on stand dynamics achieved equivalence once the selection age is reached. Although temporary and permanent increases in site quality arising from silvicultural treatments are commonly referred to as Type 1 and 2 response types, respectively (e.g., Snowdon & Waring, 1984 ), this terminology is used within a rotational context, in this study. The treatment equation modification approach has also been used to embedded genetic worth effects within empirical benchmark yield models for coniferous species in central Canada (Penner, 2004) and stand-level growth and yield simulators for black spruce and jack pine in eastern Canada (STAMAN, McInnis & Tosh, 2004 ), and for conifers in western Canada (Tree and Stand Simulator (TASS); Goudie, 2004 ).
The new response models when integrated into the algorithmic analogues of the SSDMMs were evaluated on their biological and empirical realism across a wide range of site qualities, initial densities, thinning treatments and rotation ages employing Bakuzis graphical matrices (sensu Leary, 1997 ). Based on results derived from nested simulations for each response type and species involving 5 site classes (11 - 19 m by 2 m intervals), 5 rotational lengths (50, 60, 70, 80 and 90) and 5 initial densities (1500, 2000, 2500, 3000 and 3500 stems/ha) and with and without commercial thinning treatments (35% basal area removal 25 years before rotation), indicated that the predictions from the revised SSDMMs were acceptable given the lack of evidence of consequential departures from expectation in terms of accepted stand dynamical theoretical constructs (e.g., Sukatschew’s effect, Eichhorm’s rule, self-thinning theory and yield-density relationships) and yield forecast expectations. Furthermore, the resultant stand development patterns consistently indicated an increase in temporal stand dynamics due to genetic worth effects which yielded increased volumetric and end-product outcomes and earlier stand operability status.
The site-based mean dominant height―age functions which largely govern the rate of stand development within most SDMD-based and stand-level growth and yield models, is the logical entry point for incorporating temporal developmental impacts arising from genetic worth effects. This approach is similar to the method utilized by Newton (2003) in assessing species-specific meta-based genetic worth effects for conifers in central and eastern Canada, and by Xie & Yanchuk (2003) in terms of integrating genetic worth effects within the Table Interpolation Program for Stand Yield (TIPSY) growth and yield simulator used in managing conifers in western Canada. A similar modelling approach has been used in quantifying responses to other silvicultural treatments including N-based forest fertilization (Newton & Amponsah, 2007) and chemical and mechanical vegetative management control (Newton, 2012b) . However, difference between these earlier efforts and those presented in this study relate to the response model variants considered (Type 1 and 2) and the explicit accounting for the anticipated temporal decline in the genetic gain effect over time (Lambeth, 1980) , through the use a correlative-based dissipation factor.
Plantations established with genetic improved stock for both black spruce and jack pine are relatively young (<30 yrs) and are largely without parental information in terms of genetic gain and selection age specifics. This data information gap negates an empirical assessment of the models in terms of their predictive accuracy. However, the employment of a well accepted approach (treatment modifier equation) using published genetic gain estimates derived from meta-analytical studies combined with a well-established phenotypic juvenile age-mature age correlation function, provides a conceptual basis for acceptance. Furthermore, output from a large number of simulations confirmed that the genetic worth effect models when incorporated into the SSDMM produced stand development patterns consistent with ecological axioms derived from stand dynamic and forest production theories.
Although the approach presented in this study is consistent with previous efforts used in modeling genetic worth effects, the approach is nevertheless limited given that all the genetic worth effects are assumed to be expressed through height development differences. Genetically enhanced stock for black spruce and jack pine has been propagated from 1st and 2nd generation selection strategies which have been largely based on height growth superiority (Newton, 2003) . Thus other effects, such as altered crown morphology and wood quality impacts which may accompany height growth superiority, are not accounted for in the current approach. Further research is required to determine the effect of these secondary effects on individual tree growth and overall stand dynamics before the full extent of genetic worth effects on forest productivity is known with certainty.
5. Conclusion
The analytical approach used to account for the accelerated rate of stand development arising from genetic worth effects was through adjustments to the site-based dominant height-age functions given that these functions govern stand dynamics and structural change within SSDMMs. The presented models are also applicable to past generations of the SDMD-based models and other height-driven growth and yield simulators used in forest management planning. Conceptually, the approach parallels the analytical logic proposed by others for projecting genetic worth effects and falls within the treatment modifier equation class of modeling methodology used to quantify responses to silvicultural interventions. The results of this study provide additional support for this approach in that the development of species-specific modified height-age functions and embedding them into stand-level decision support models enabled greater predictive realism in long-term yield forecasts. Although empirical predictive ability was not explicitly addressed, the revised models predicted stand development patterns that were consistent with axioms of even-aged stand development.