Comparison of temperature preference for habitat among three species of oceanic sea skaters, Halobates micans, H. germanus and H. sericeus ()
1. INTRODUCTION
Many ectotherms of diverse species modify the time of day for conducting behaviors (such as using sun or shade) [1-5], and maintaining body temperature within a narrow, species-specific range, at least during their active time [6-9]. Temperature preferences shown by insects have adaptive meaning for their life. A study on the temperature preference of grain beetles showed that they prefer warmer areas, and increasing the temperature caused a greater number of beetles to move towards the warmer areas (Cryptolestes ferrugineus: [10]; Liposcelis: [11]). Preference for warmer places can be advantageous for inducing a rapid increase in population number in the grain stock.
Differences in temperature preference could play an important role in habitat variability in some cases. One example is the relationship between benthic insects (Ephemeroptera, Plecoptera, Coleoptera, Trichoptera) and temperature, studied in small and medium-sized streams in Germany [12]. Up to 29% of the variability in the Ephemeroptera, Plecoptera, Trichoptera and Coleoptera, communities were explained by summer temperature variation in the data sets for both small and mediumsized streams.
From an eco-physiological point of view, the temperature preference of animals could be related to the ambient temperature of their habitats. Girguis and Lee [13] investigated adaptation to extreme temperature by examining the temperature preferences of the polychaete worm species, Alvinella pompejana and Paralvinella sulfincola (family Alvinellidae), living directly on hightemperature vents in the Eastern and Northeastern Pacific, respectively, where temperatures up to 81˚C and 90˚C have been measured around A. pompejana and P. sulfincola tube openings, respectively [14]. P. sulfincola worms were shown to be thermo-taxic and extremely thermo-tolerant, preferring temperatures between 40˚C and 50˚C, with individuals surviving 7 hours of chronic exposure to 50˚C and 15 minutes of acute exposure to 55˚C.
Few studies have examined the physiological mechanism of temperature preference in insects. One study showed that dopamine signaling in mushroom bodies of the brain regulated temperature preference behavior in Drosophila flies [15]. Various genetic studies were performed to identify the functions of dopamine in temperature preference behavior. Inhibition of dopamine biosynthesis by genetic mutation or treatment with chemical inhibitors induced flies to prefer colder temperatures than normal.
Oceanic sea skaters inhabit the open ocean where the sea surface temperature is very stable, at around 30˚C in tropical areas throughout the year [16,17]. However, the temperature preference of the three species of oceanic sea skaters, Halobates micans (Body size: 4.9 - 5.1 mm [16]), H. germanus (3.4 - 3.6 mm) and H. sericeus (3.2 - 3.4 mm), in colonizing temperate and subtropical ocean areas, has yet to be determined. During one cruise by the R/V Hakuhomaru in May, 2003, one sea skater species, Halobates sericeus, was collected at 18 locations sampled in the East China Sea (27˚10ʹN - 33˚24ʹN, 124˚57ʹE - 129˚30ʹE) [18], and H. micansand/or H. germanus at 8 locations in the area south of 29˚47ʹN, where the water temperature was higher than 25˚C. Samples collected at three locations where the water temperature was less than 23˚C did not contain either H. micans or H. germanus. In a pilot experiment, Harada et al. [19] showed decreasing of heat tolerance in the order of H. micans, H. germanus, then H. sericeus, suggesting that the two preceding species were likely to prefer higher temperatures for their habitat than H. sericeus. However, data in other oceanic areas are needed to support this hypothesis on diversity of temperature preference among the three species. We thus set out to examine the relationship between population density, water surface and air temperature in the three species of oceanic sea skaters, Halobates micans, H. germanus and H. sericeus, inhabiting tropical and subtropical-temperate areas of the Pacific Ocean.
2. MATERIALS AND METHODS
Samplings were taken with a Neuston net (6 m long and with diameter of 1.3 m) during the MR-12-05 cruise (The fifth cruise in 2012 by Research Vessel MIRAI) (Figure 1) by the Research Vessel, MIRAI (government vessel for oceanography: 8687 t) and KH-13-02 (The second cruise in 2013 by Research Vessel, HAKUHOMARU (government vessel for marine science: 3991 t) (Figure 2), both owned by the Japan Agency for Marineearth Science and TECHnology (JAMSTEC), and with an ORI net (6 m long and with diameter of 1.5 m) during the MR-06-05-Leg 3 cruise (Figure 3), also by the R/V MIRAI. Samplings were taken on November 12, 15 and 22, 2012 during the MR-12-05, from February 4 to March 7, 2013 during the KH-13-02, and from December 24, 2006 to January 8, 2007 during the MR-06-05- Leg 3.
The net was trailed for 15 minutes, 3 - 6 times per station exclusively at night with a ship speed of 2.0 - 3.0 knots to the sea water. The surface area swept by Neuston (ORI) net was evaluated as an expression of flowmeter value × 1.3 m (or 1.5 m) of the width of the net. The population density was calculated as individual number per 1 km2 using the expression, (1,000,000/surface area swept by the net) × individual number of oceanic sea skaters collected.
Samples were transferred from a sample bottle at the end of the net to a transparent aquarium. All the samples were transferred with tweezers used for plankton from the transparent aquarium to a piece of white paper on a tray. All specimens were examined very carefully to identify species and stage (according to Andersen and Cheng [16]) and were tallied. The temperature of the surface water and air temperature on the ship deck were measured at all samplings.
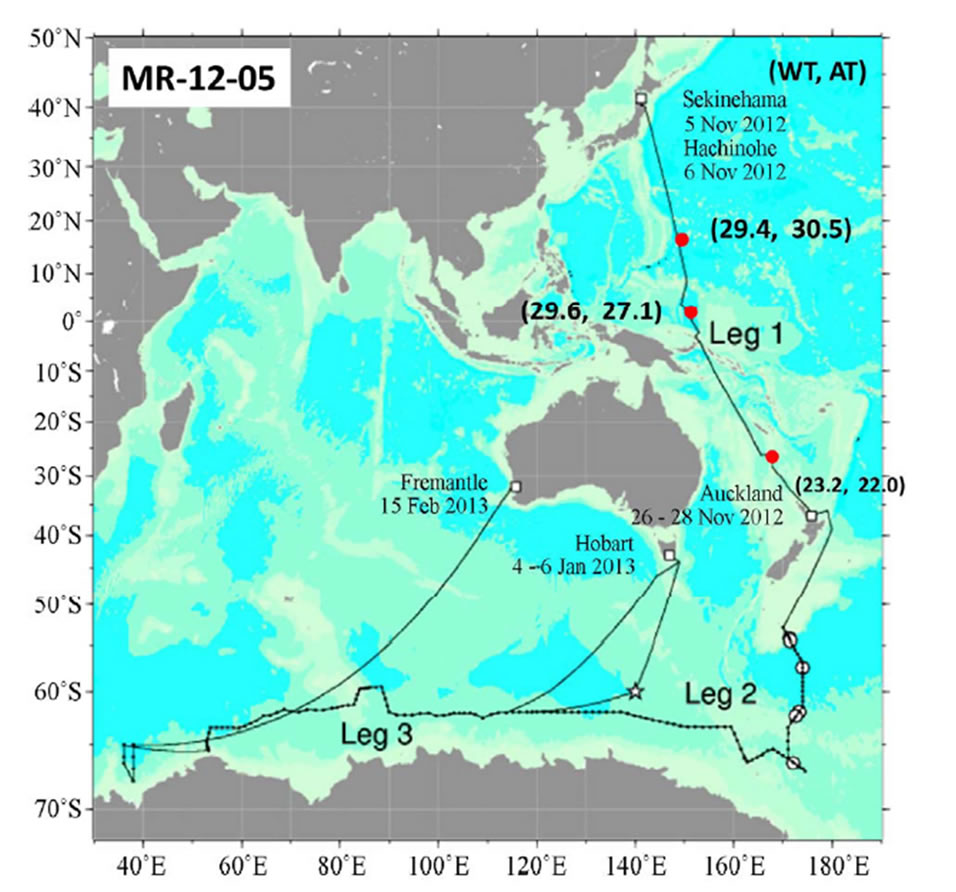
Figure 1. Cruise track and sea skater sampling points (red circles) during the no: MR-12-05 by the R/V MIRAI.
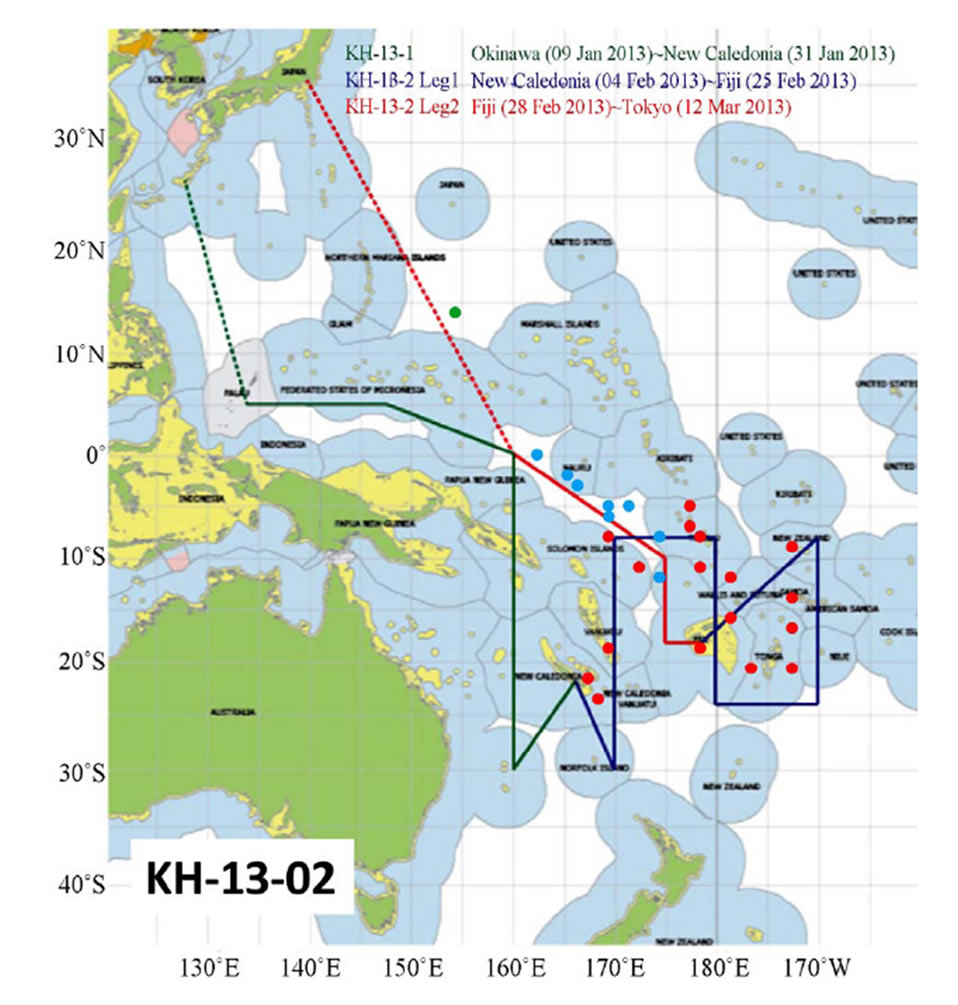
Figure 2. Cruise track and sea skater sampling points during cruise no: KH-13-02 by the R/V HAKUHOMARU. Sampling points are shown by red circles in relatively higher latitude areas in the southern hemisphere during KH-13-02-Leg 1 and blue circles in relatively lower latitude areas during the KH- 13-02-Leg 2. One isolated sampling point at 15˚N during the KH-13-02-Leg 2 is shown by a green circle.
3. RESULTS
3.1. Population Density
In the tropical to temperate zones of the central Pacific Ocean with a longitude of 150˚ - 165˚E, the smallest species, Halobates sericeus, was dominant at 14˚N and 27˚S, while H. germanus, medium in size among the three species, was dominant at the low latitude of 2˚N (Table 1). In the southern hemisphere of the Pacific Ocean, H. micans, the largest species, was dominant in the relatively lower latitude of 0˚ - 10˚S during the KH-13-02- Leg 2, while H. germanus was dominant in the higher latitude of 10˚ - 25˚S during the KH-13-02-Leg 1 (χ2- test: χ2-value = 112.4, df = 1, p < 0.001) (Tables 2A and B). H. sericeus was present at a high density of 40,000 individuals/km2 at 15˚N, 152˚E (Table 2C). In the western tropical Pacific Ocean at 0˚ - 8˚N, 130˚ - 135˚E, both H. micans and H. germanus were present in high densities during the MR-06-05-Leg 3 (Table 3).
3.2. Temperature and Population Density
Among the three sampling points, H. sericeus, H. germanus and H. micans were present in the highest density at points with temperatures of 22˚C - 23˚C, 27˚C - 29˚C, and 29˚C - 31˚C, respectively (Figures 4 and 5).
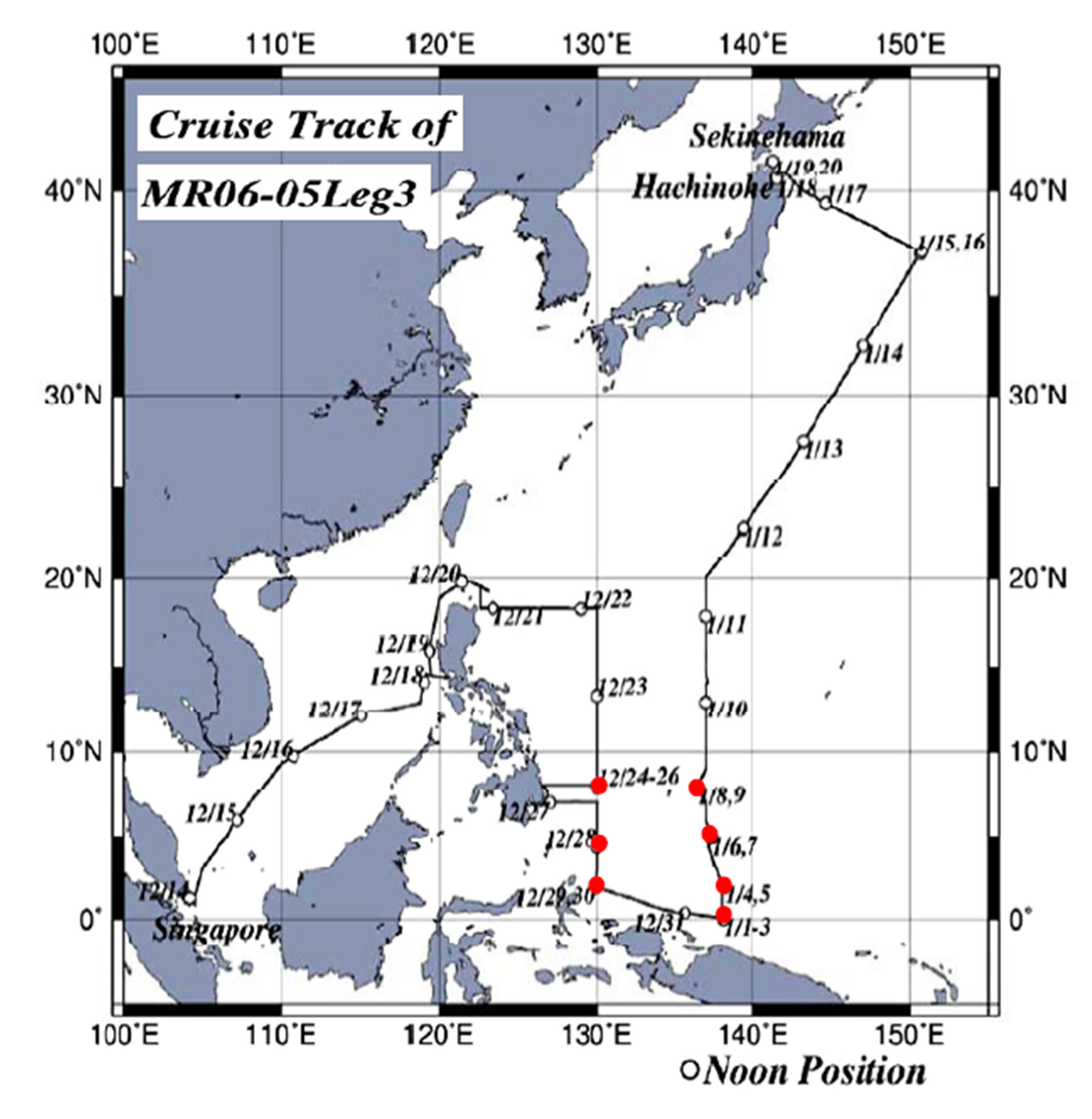
Figure 3. Cruise track and sea skater sampling points (red circles) during the no: MR-06-05-Leg by the R/V MIRAI.
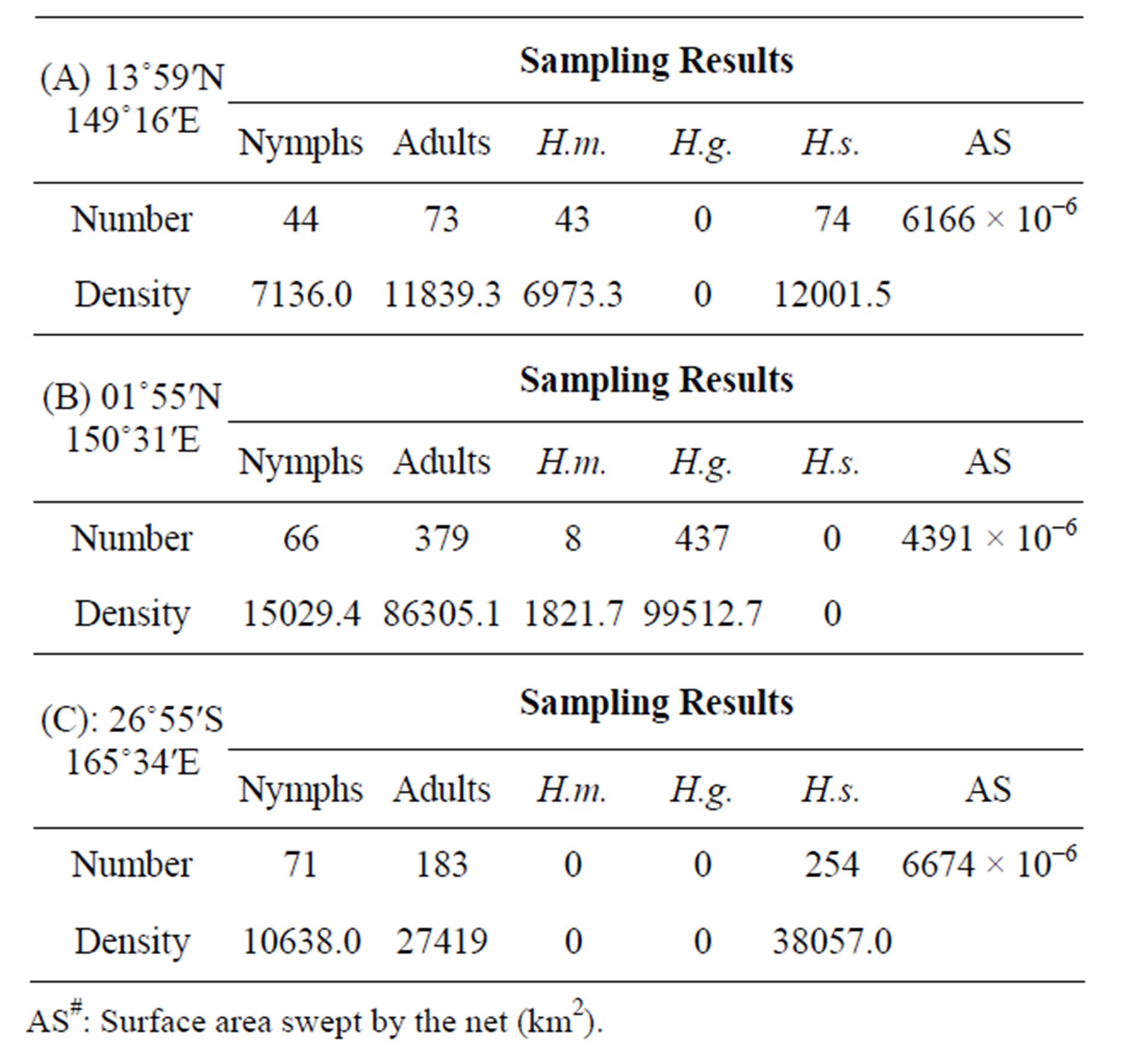
Table 1. A comparison of population density of oceanic sea skaters, Halobates among five areas of the open Pacific Oceans. Samplings were performed during five cruises, including this cruise. H.m: Halobates micans; H.g.: H. germanus; H.s.: H. sericeus; H.p.: H. princeps; sp.: H. sp.; Density: individual number/km2; MR-12-05-Leg 1 (Stations 1-3: A-C): Western Subtropical and Tropical Pacific Ocean.
In the tropical to temperate zone of the southern hemisphere of the Pacific Ocean, both species and temperature of air and water tended to be related to population density independently based on ANCOVA (Analysis of CO-Variance: air temperature on density with covariance
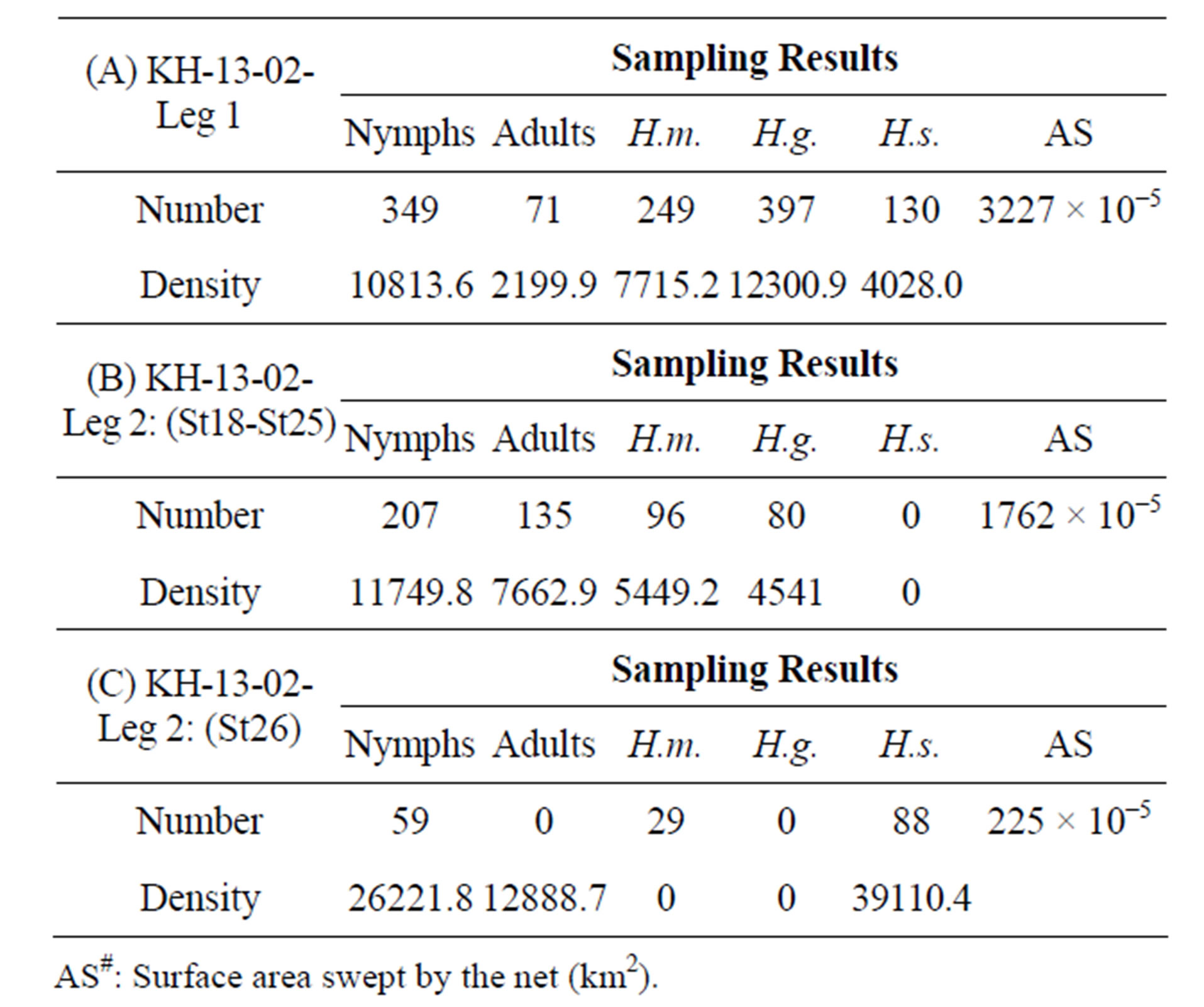
Table 2. A comparison of population density of oceanic sea skaters, Halobates among five areas of the open Indian and Pacific Oceans. Samplings were performed during five cruises, including this cruise. H.m: Halobates micans; H.g.: H. germanus; H.s.: H. sericeus; H.p.: H. princeps; sp.: H. sp.; Density: individual number/km2; KH-13-02-Leg 1-2 ((A)-(C)).
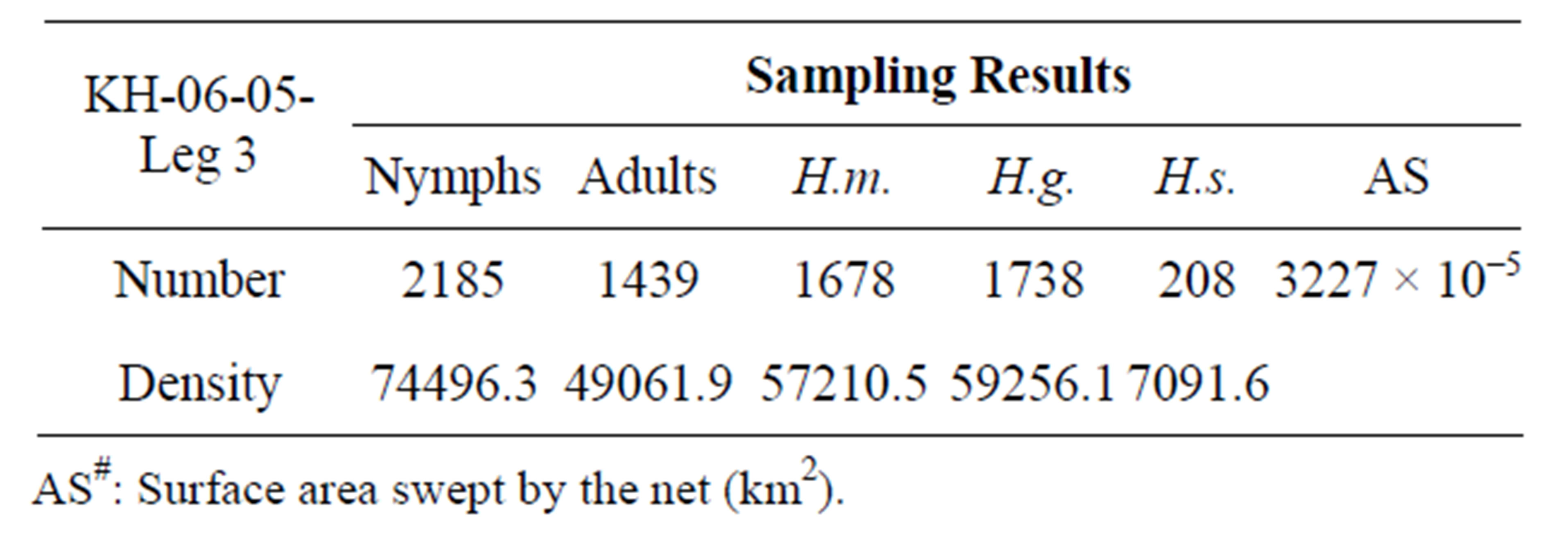
Table 3. A comparison of population density of oceanic sea skaters, Halobates among five areas of the open Indian and Pacific Oceans. Samplings were performed during five cruises, including this cruise. H.m: Halobates micans; H.g.: H. germanus; H.s.: H. sericeus; H.p.: H. princeps; sp.: H. sp.; Density: individual number/km2; MR-06-05-Leg 3 (Stations 1-7): Western Tropical Pacific Ocean (0˚ - 8˚N, 130˚ - 138˚E).
of species, df = 1, F = 2.46, p = 0.093; water temperature on density with covariance of species, df = 1, F = 2.46, p = 0.093; species on density with covariance of air and water temperature, df = 3, F = 5.55, p = 0.002).
Positive and negative correlations were seen between population density and both air and surface water temperature for H. micans and H. sericeus, respectively, but no correlation was seen for H. germanus in the tropical and temperate areas of mainly the southern hemisphere in the Pacific Ocean (Figures 6 and 7). In the western tropical Pacific Ocean, the only association seen was a significant positive correlation between population density and air and water temperature for H. germanus (Figures 8 and 9).
4. DISCUSSION
Cheng [20] stated that the occurrence and abundance of oceanic sea skaters were primarily controlled by surface sea water temperature. The optimal temperature for oceanic sea skaters was estimated to be 24˚C - 30˚C [20,21]. However, Andersen and Cheng [16] claimed the lower limit of surface water temperature for survival of oceanic sea skaters to be around 25˚C.
The results of the present study suggest that temperature preference by oceanic sea skaters differs depending on the species. Halobates sericeus may prefer a lower range of temperatures at around 23˚C - 28˚C than the other two species, H. micans and H. germanus, that favor a relatively higher range of temperatures, at around 28˚C - 30˚C.
The lower and wider-ranging values for temperature preference for habitat by H. sericeus may relate to its wide-ranging latitudinal distribution from 5˚N - 42˚N [17], across which the Northern Equator Current and the Kuroshio (Black Current) may transfer this species from lower to higher latitudes, while the latitudinal habitat
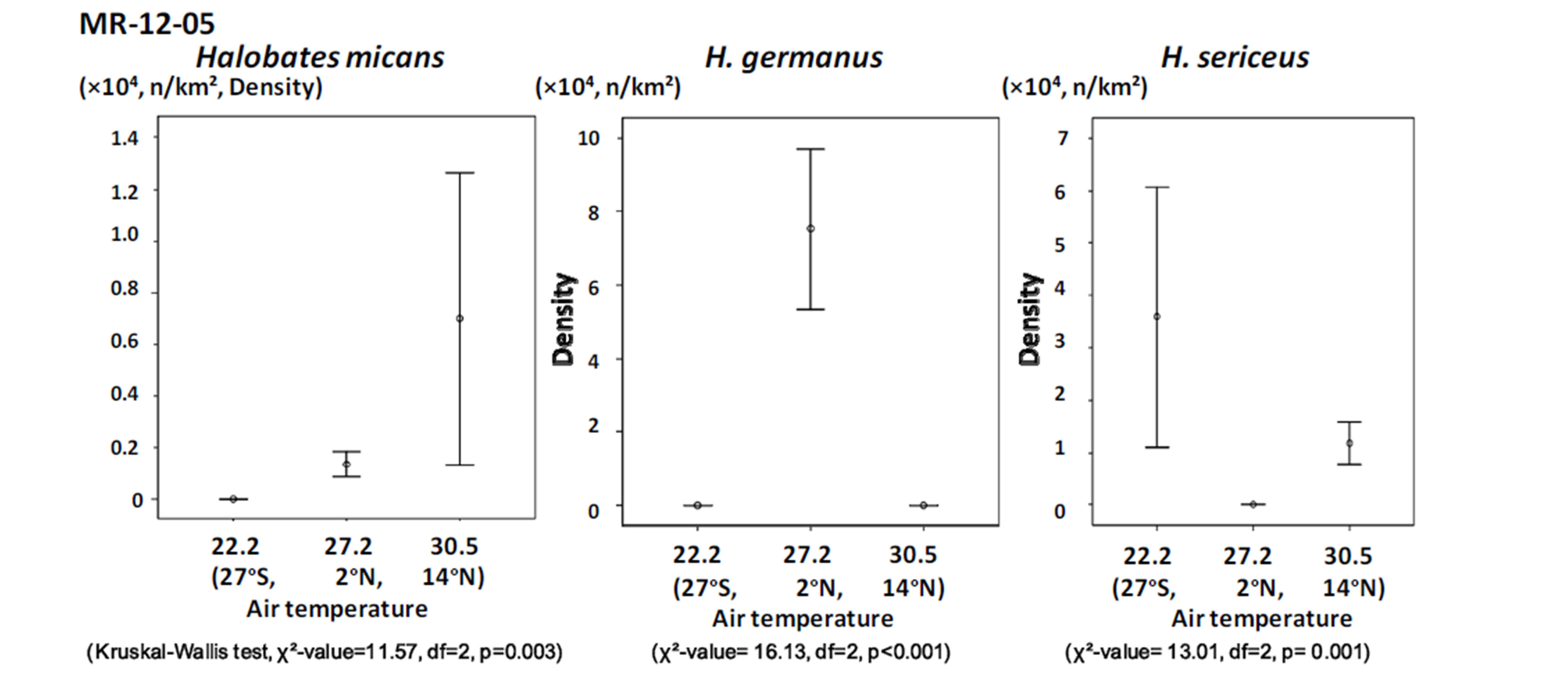
Figure 4. The relationship between population density of the three species of oceanic sea skaters and air temperature in the three sampling points during the cruise MR-12-05.
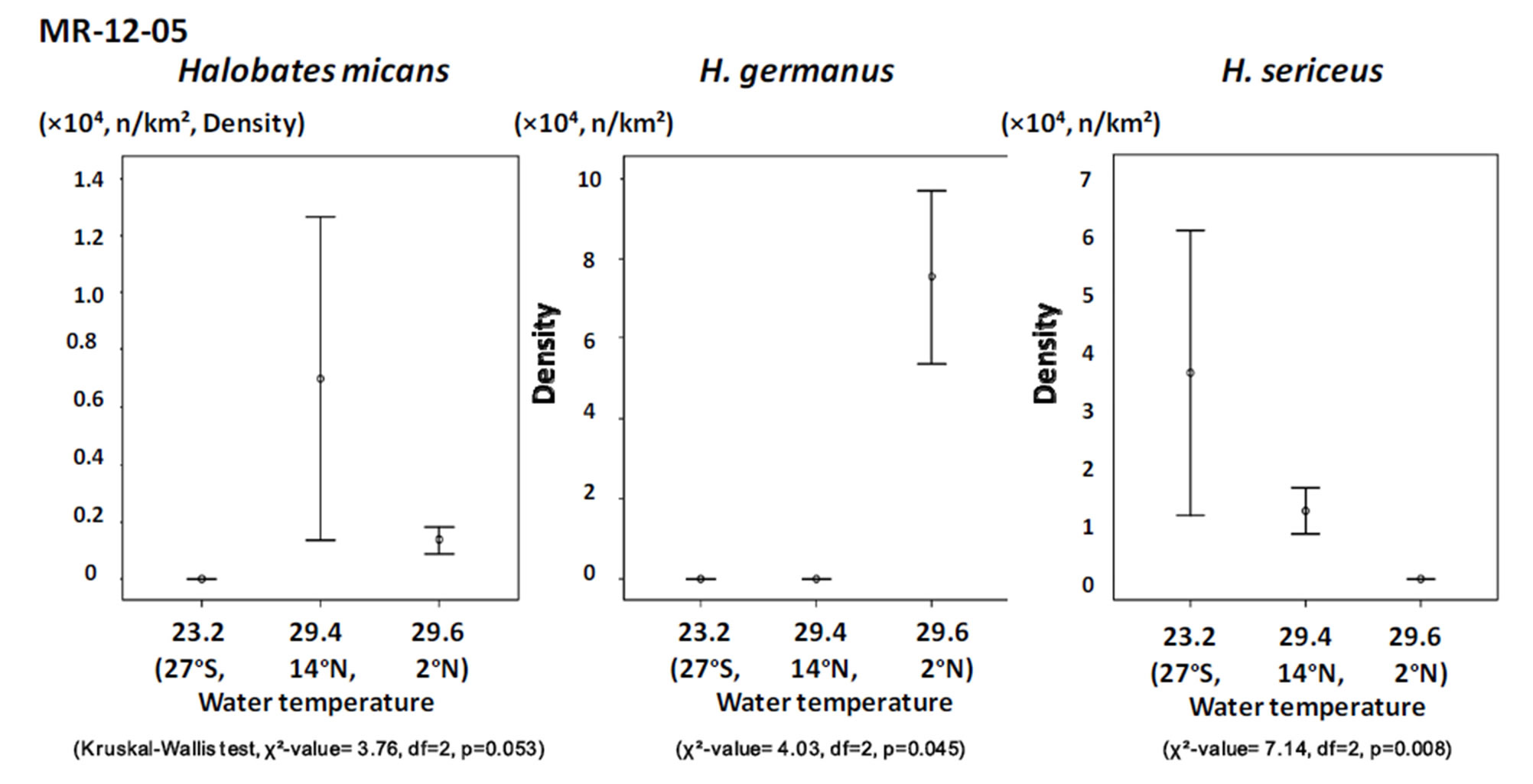
Figure 5. The relationship between population density of the three species of oceanic sea skaters and surface water temperature in the three sampling points during the cruise MR-12-05.
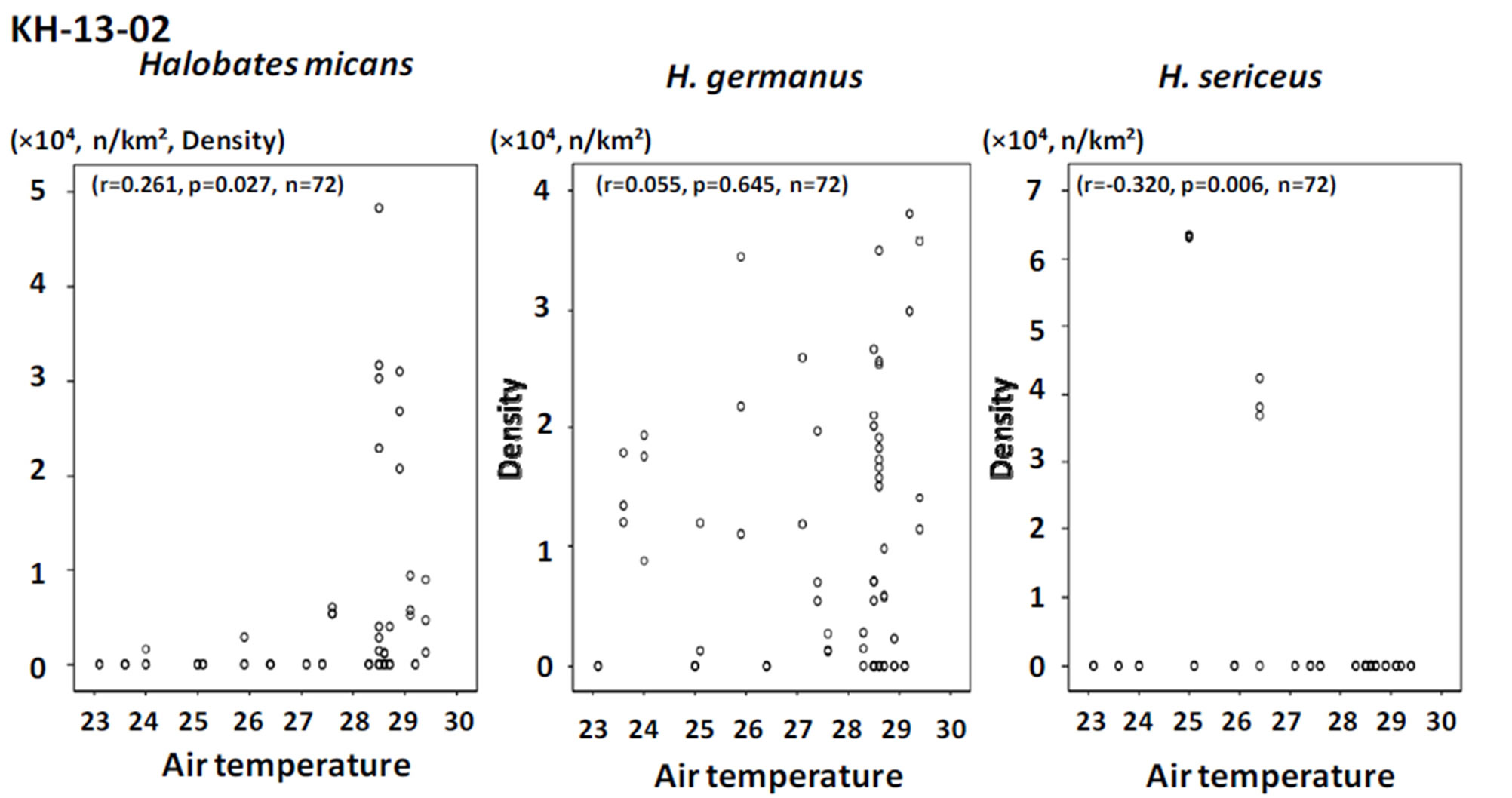
Figure 6. The relationship between population density of the three species of oceanic sea skaters and air temperature in the sampling points during the cruise KH-13-02.
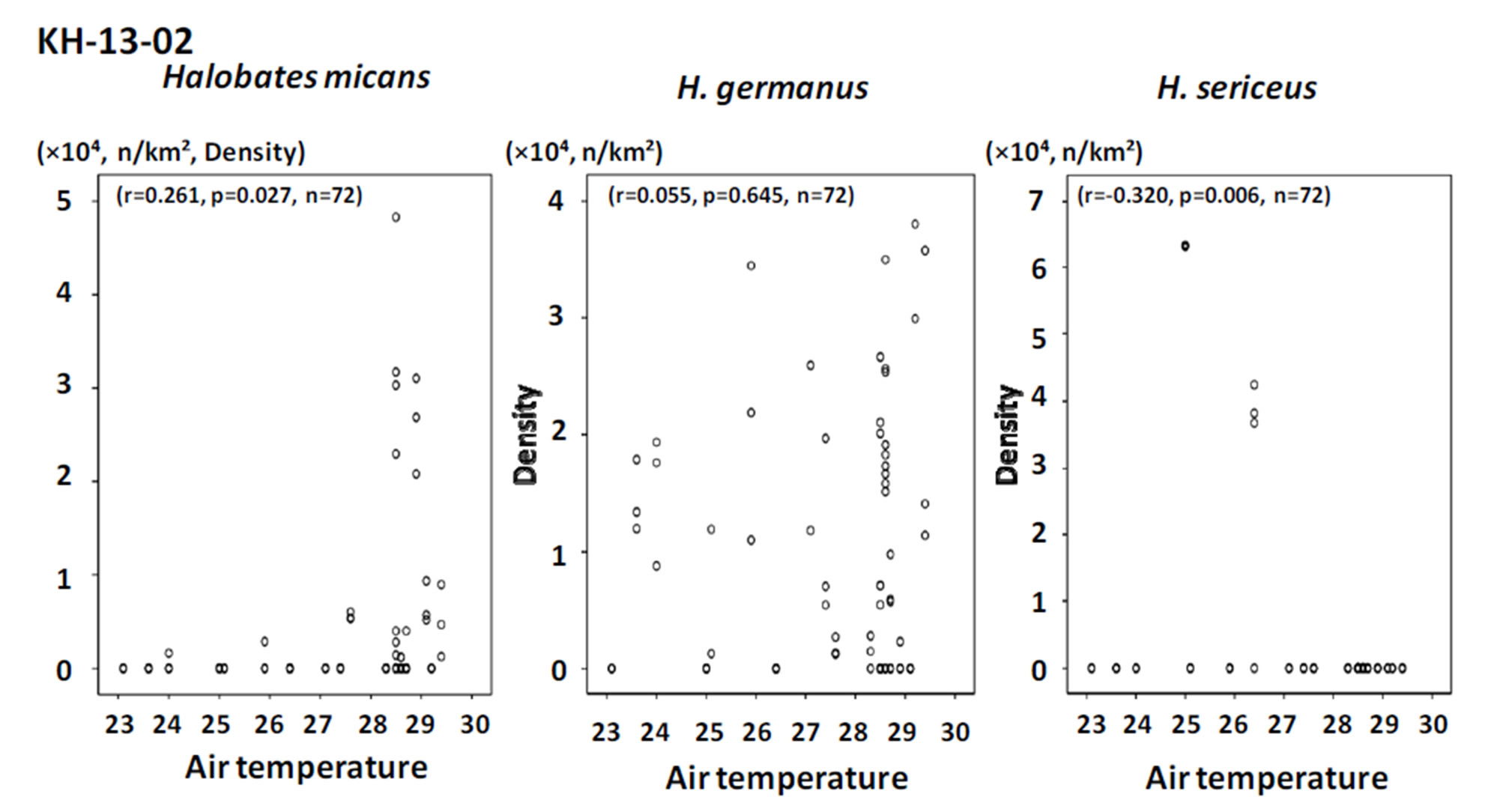
Figure 7. The relationship between population density of the three species of oceanic sea skaters and surface water temperature in the sampling points during the cruise KH-13-02.
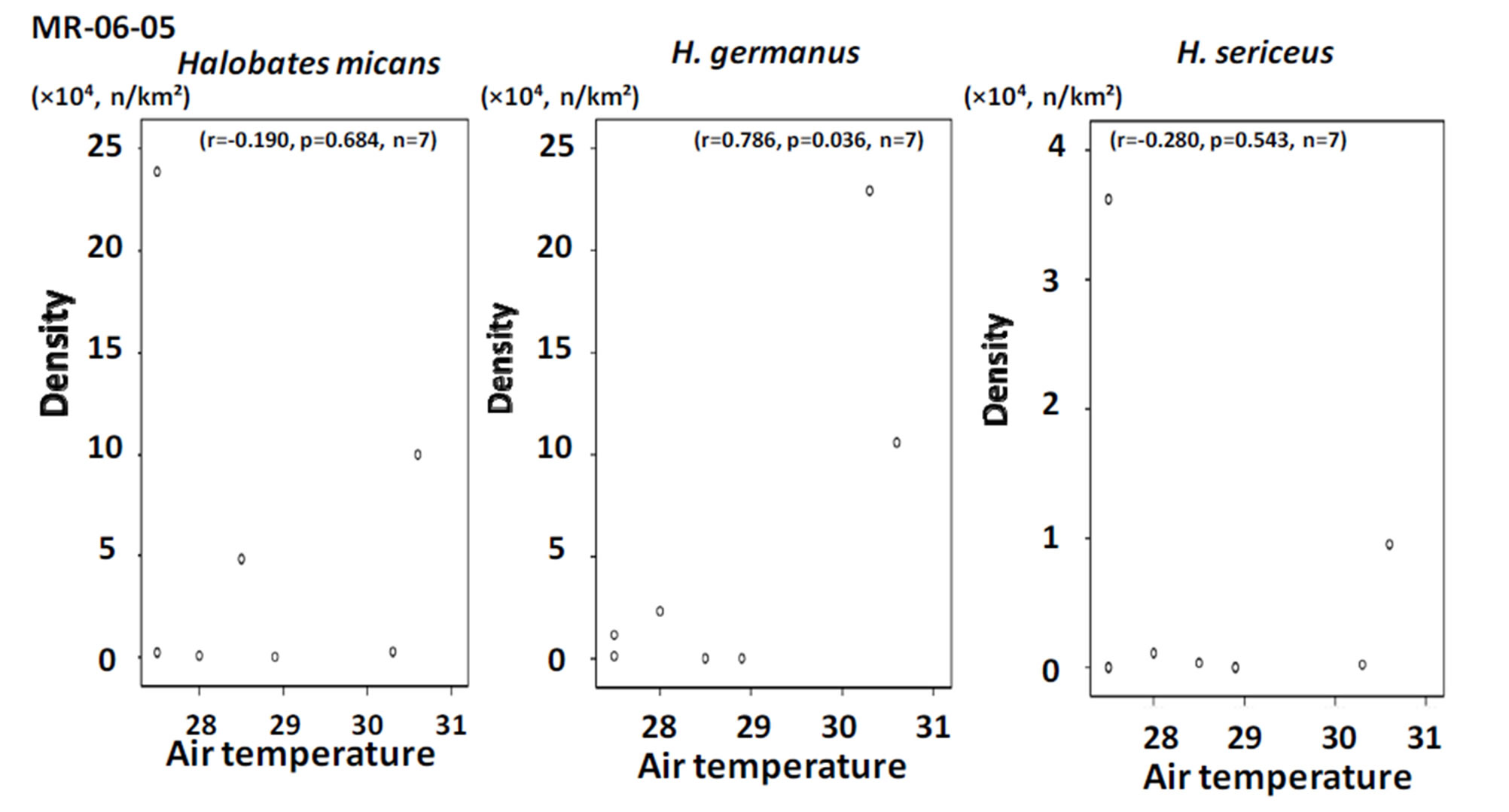
Figure 8. The relationship between population density and air temperature in the sampling points during the cruise MR-06-05.
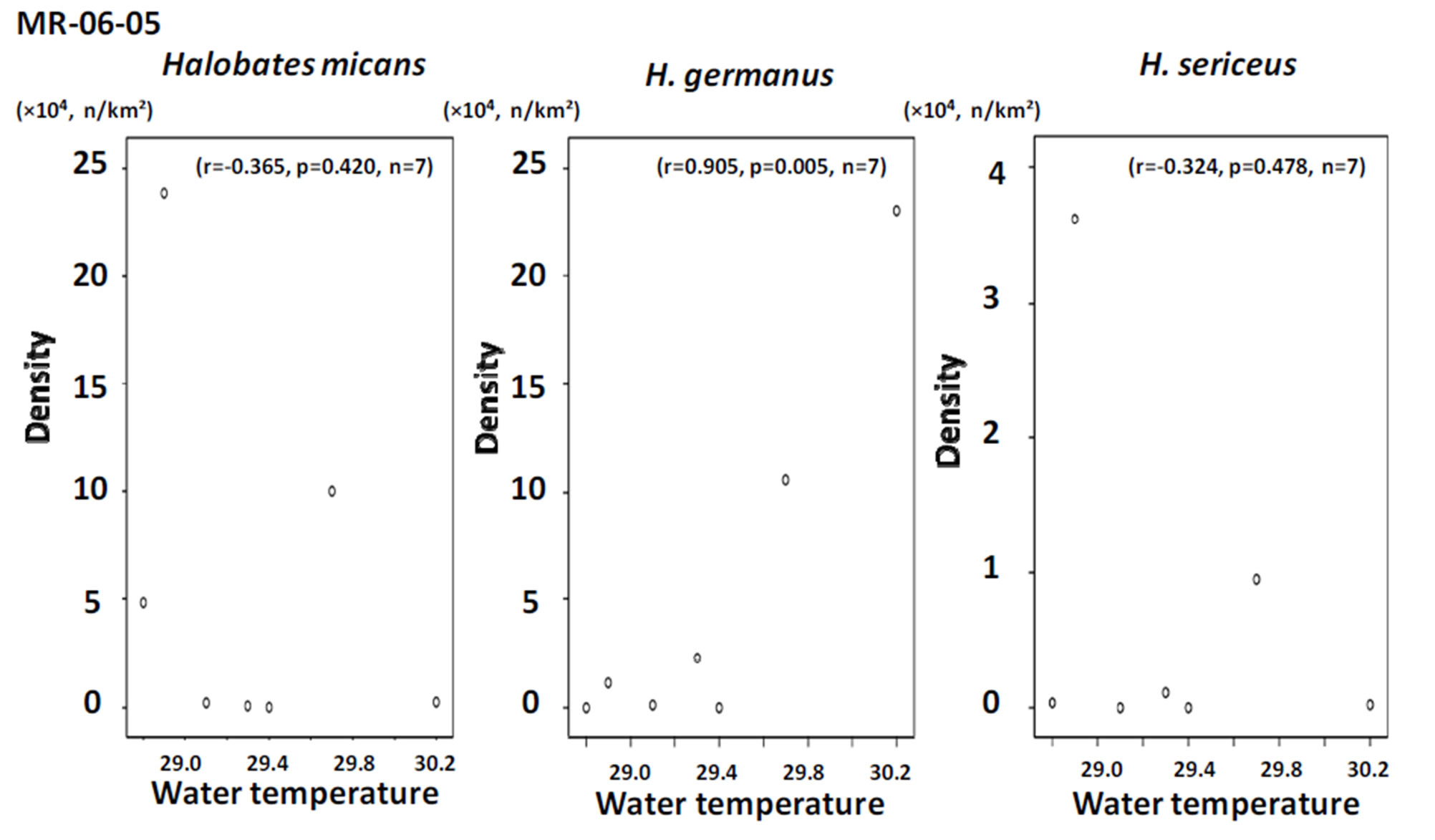
Figure 9. The relationship between population density and surface water temperature in the sampling points during the cruise MR-06-05.
range shown by H. micans and H. germanus is much narrower, mainly from 15˚S to 15˚N [17,19,22]. The wide range of temperature preference shown by H. sericeus may be advantageous for its distribution over a wide range of latitudes.
Alternatively, the preference for lower temperature by H. sericeus could be related to its size as the smallest among the three species, giving it less competitive power for occupying an adequate area at high temperatures for growth and reproduction. If the global warming will be going on in the future, northern spreading of the distribution of H. micans might be speculated in accordance with the warmed surface water.
As a limitation of this study, no experimental data were obtained on the effects of temperature on life history traits (such as fecundity, survival rate of larvae and adults, frequency of mating behavior, and hatching rate of the first instars). Further studies are needed to obtain such experimental data.
5. ACKNOWLEDGEMENTS
We would like to thank Dr. Yuji KASHINO (Head Scientist on Leg 3 of the cruise: MR-06-05, JAMSTEC), Dr. Katsuro KATSUMATA (Head Scientist of the cruise: MR-12-05, JAMSTEC) and Prof. Jun AOYAMA (Chief Scientist of the cruise: KH-13-02-Leg 2, University of Tokyo) for their permission to do this study during the cruises on the R/V MIRAI and R/V HAKUHOMARU, their warm suggestions on this study, and their encouragement and help throughout these cruises. The field studies including samplings during the three cruises were also possible due to support from all of the crew (Captains: Mr. Yuji KITA for MR-06-05-Leg 3, Mr. Yoshiharu TSUTSUMI for MR-12-05 and Mr. Takatoshi SEINO for KH-13-02) and all the scientists and engineers from Global Ocean Development Inc. (GODI) and Marine Work Japan (MWJ) in these cruises. We would like to give them special thanks. Thanks are also due to Ms. Laura SATO for her professional English editorial work on this manuscript.