Infraspecific Delimitation of Acacia senegal (Fabaceae) in Uganda351
bark and dark pink underbark. The branches are domi-
nantly straggling with a dark pink underbark colour
while the branchlets are mainly papery and peeling with
some individuals peeling but not papery. Leaves are
dominantly prickly with leaf breadth (1.43 - 6 cm), pinna
length (0.86 - 2.1 cm), number of leaflet pairs (10 - 14
{16}) and number of pinna pairs (3 - 4). The taxon bears
characteristically more oblong than circular petiolar
glands of length (4.0 - 7.4 µm) and breadth (2 - 3.4 µm)
while the rachis glands length varies from 3 to 4.4 µm
and breadth from 2 to 2.8 µm. The taxon therefore has
petiolar glands whose length to breadth ratio is ≥ 2.0
making the glands characteristically more oblong than
circular. The calyx is purplish-green to red and the pods
bear mainly rounded apical shapes.
This description fits and even improves the previous
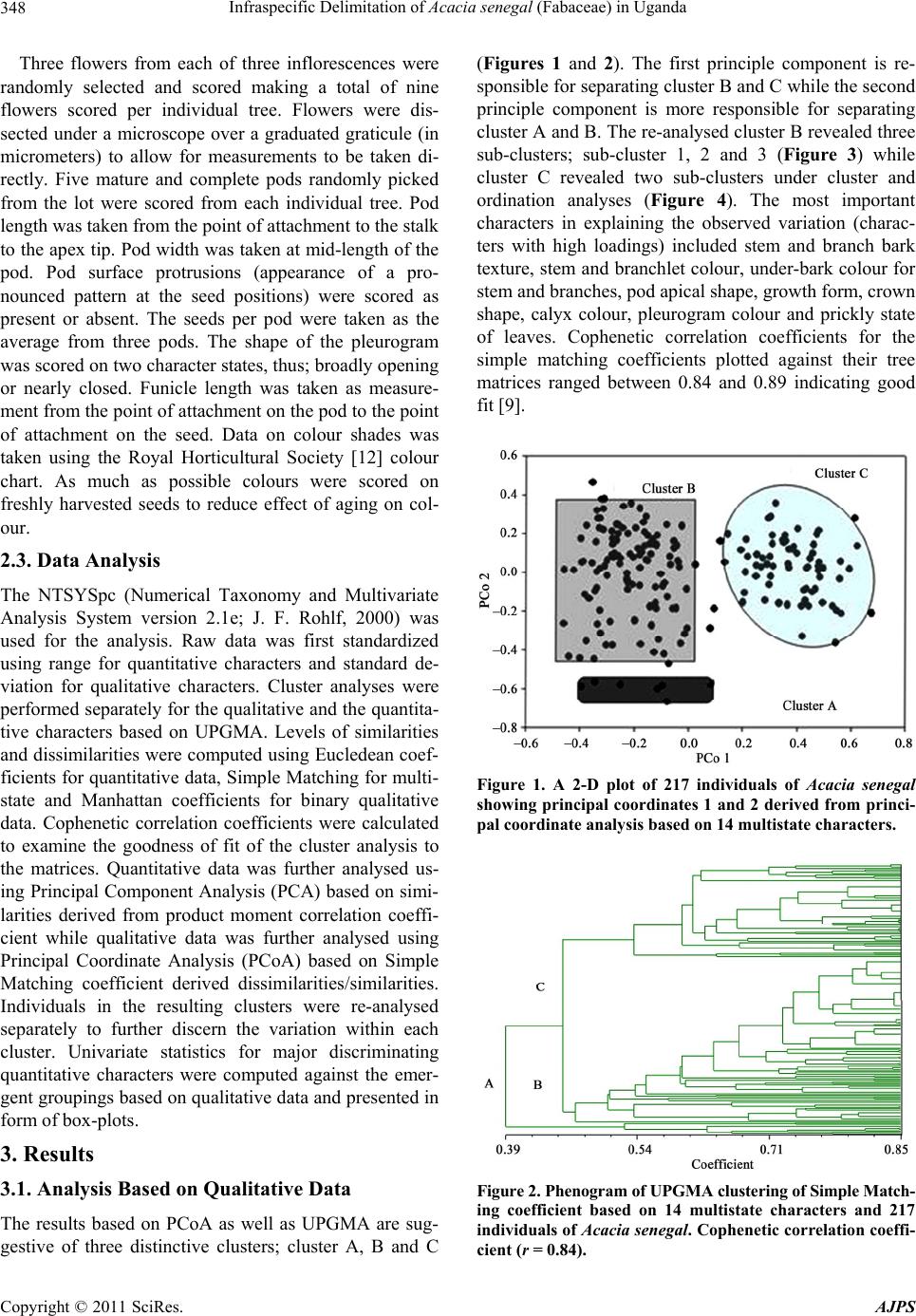
descriptions of the variety kerensis [4,7]. Figure 1 also
clearly brings out the closer relationship between this
variety and var. senegal (cluster B) which could have led
to the conclusion by [11] that the two varieties blend into
each other and are therefore one variety.
The variety senegal is here described as mainly a tree
with some shrubs, (1.8 - 7.1 m ht.) with a very variable
crown shape (lax-rounded, flat spreading to open irregular).
The stem bark is papery and peeling to fissured, green-
yellow, but also grey-brown, dull grey and occasional
bright orange brown, while the underbark is cream-white,
dark pink but predominantly dark pink. The branches are
straight to straggling, papery and peeling while the young
branchlets are yellowish to purplish black. The leaves are
seldom prickly, with a leaf breadth of 2.1 - 3.8 cm, pinna
length of 1.3 - 2 cm, number of leaflet pairs 12 - 15 (16)
and number of pinna pairs (3 - 5). The petiolar gland
length varies from 4.0 to 5.8 µm while the breadth varies
between 2.0 and 3.2 µm. The rachis gland length is 3.4 -
4.4 µm while the breadth is 2 - 3.2 µm. The calyx is pur-
plish-green and the pods apices acute to rounded. This
description agrees with and further augments the findings
of [7,11]. Neither the degree of pubescence on the bran-
chlets nor on the inflorescence axis, which were used
ambiguously by earlier studies [14] to separate the varie-
ties, has been found to be important in this study. No
wonder [3] retracted this position and later conceded [4]
together with [5] that the taxonomy of the species was far
from clear. This study has therefore provided better de-
limitation of the taxon significantly removing previous
contradictions.
From these findings, Acacia senegal var. leiorhachis
can be described as a tree (4.2 - 13.9 m ht.) with a lax-
rounded to flat spreading crown. The stem bark is fis-
sured, grey-brown to dull grey whereas the underbark is
red. The branches are straight to straggling, purplish-grey
to purplish-black, papery and peeling, peeling and not
papery, to papery and not peeling with a red underbark.
The leaves are seldom prickly with leaf breadth (2.7 - 5.6
cm), pinna length (1.7 - 3.26 cm), number of leaflet pairs
(16 - 20) and pinna pairs (4 - 5). The petiolar gland
length varies between 2.4 and 5.4 µm while the breadth
varies between 2.4 and 5 µm and characteristically more
circular than oblong. The rachis gland length varies be-
tween 2.6 and 4.6 µm and the breadth from 2.0 and 4.2
µm. The calyx is purplish green while the pod apical
shape is acute to round. The description agrees with and
complements that of [7] together with the arguments
thereof. Due to the absence of sufficiently distinguishing
characters on herbarium specimens, [15] decided that the
differences between A. senegal and A. circummarginata
were not distinct and merged the two into A. leiorhachis.
The present study has identified characters that can be
used to distinguish the different taxa in herbaria (as well
as in the field) thus agreeing with [5,11] that the taxa are
distinct. However, [11] preferred to maintain this taxon
as a species (A. circummarginata) following [3] as op-
posed to a variety (leiorhachis) as given by [5] and sub-
sequently followed by [3]. Our opinion based on the
findings of this study is that this taxon (var. leiorhachis)
as well as var. senegal are better treated as varieties of A.
senegal.
The second level of taxonomic differentiation delim-
ited three groups from var. senegal (Figure 3) and two
from var. leiorhachis (Figure 4). The three groups de-
limited from var. senegal can be described as follows.
The first group, represented as cluster 1 (Figure 3), pre-
sents as a tree with a lax-round crown, fissured green-
yellow stem bark, yellowish papery and peeling bran-
chlets, purplish-green calyx and round pod apices. The
second group, represented as cluster 2, differs from the
first one by having dull-grey to grey-brown, papery and
peeling stem bark, purplish-black branchlets, and acute to
acuminate pod apices. This group was most dominant.
The third group, represented as cluster 3, presents as a
shrub with flat spreading to open irregular crown.
The two groups delimited from var. leiorhachis (Fig-
ure 4) can be described as follows; the first presents as a
tree with grey-brown stem bark and mainly straight
branches while the second one presents as a tree with
dull-grey stem bark and straggling branches.
The groups delimited at the second level of differen-
tiation can be based described as variants of variety
senegal and leiorhachis respectively agreeing with [6]
that varieties of Acacia senegal can develop into differ-
ent growth forms. A taxonomic key is hereby presented
but since the individuals studied were from only Uganda,
the key cannot be treated as a universal one until similar
studies are done across the species range of occurrence.
The ecological significance of this differentiation will be
further investigated in our subsequent study.
Copyright © 2011 SciRes. AJPS